![]()
![]()
Utskrift 07.07.2026 06:41
![]()
![]()
Utskrift 07.07.2026 06:41
Gjennom prosjektet «Plantevernhåndbok for økologisk landbruk» har vi skrevet 4 bøker med felles tittel «Plantevern og plantehelse i økologisk landbruk»:
I tillegg til å kunne kjøpes i bokformat vil de 4 bindene også finnes i nettversjon på «Agropub» (http://www.agropub.no). Primær målgruppe for bøkene er bønder og hagebrukere, men gjennom bruk av såkalte «tekstbokser» med fordypningsstoff tror vi også at studenter på videregående trinn og ved høyskoler vil ha stort utbytte av stoffet som vi presenterer. Mens Bind 1 er ganske grunnleggende i sin oppbygning, er de 3 siste bindene betydelig mer anvendt og mer en «kokebok» for hvordan man best kan håndtere de ulike skadegjørere.
I Bind 1 går vi relativt grundig til verks, ikke minst gjelder dette kapittel 2 om de ulike skadegjøreres livsstrategier. Kunnskap om skadegjørerens biologi og økologi er viktig både for i forkant å unngå at plantevernproblemer oppstår, og for å gjennomføre direkte mottiltak i kulturene på en best mulig måte. Det å «kjenne fiendens» sterke og svake sider er svært viktig for å løse et aktuelt plantevernproblem, men vi tror at slik kunnskap rett og slett også gjør det morsommere å være plantedyrker. Mange skadegjørere har jo en ganske så finurlig biologi og sameksistens med sine omgivelser for å sikre egen vekst og «videreføring av slekta» gjennom produksjon av avkom. Vi håper dessuten at kunnskapen i disse bøkene skal være med å stimulere til mer robuste dyrkingssystemer hvor forebyggende plantevern har en sentral plass. Hvis framgangen til økologisk landbruk skal fortsette er det dessuten svært viktig at plantevern- tiltak og strategier harmonerer med målsetningene for økologisk landbruk, ikke minst gjelder dette mht miljøkonsekvensene. Vi kan dessverre ikke love at vi for alle skadegjørere har effektive og gode ikke-kjemiske råd, men med en fortsatt satsing på forskning tror vi at det i framtidige versjoner av «Plantevernhåndboka» også vil komme gode løsninger for disse.
Bioforsk Plantehelse og Bioforsk Økologisk har vært formelle samarbeidspartnere gjennom hele prosjektperioden, men Høyskolen i Hedmark har også vært en viktig bidragsyter. Redaksjonen til «Plantevernhåndboka» har i tillegg til undertegnede bestått av Svein Magne Birkenes, Birgitte Henriksen og Richard Meadow som alle arbeider ved Bioforsk Plantehelse, og Theo Ruissen som er ansatt ved Bioforsk Økologisk.
I tillegg til redaksjonen har også mange andre stått sentralt i skrivearbeidet av Bind 1: Helge Sjursen (Bioforsk Plantehelse) i om «Ugrasets livsstrategier» (Kap. 2), Reidar Holmøy (pensjonist, tidligere UMB) om teknisk utstyr for ugraskontroll (Kap.4), Kjell Mangerud (Høyskolen i Hedmark) om jordarbeiding (Kap.3) og teknisk utstyr (Kap.4). Ricardo Holgado og Bonsak Hammeraas (Bioforsk Plantehelse) om nematoder som skadedyr.
Følgende personale fra Bioforsk Plantehelse: Linnea Wang og Jan Netland, Gunn Mari Strømeng og Linda Gordon Hjeljord, samt Ingeborg Klingen, Solveig Haukeland og Trond Hofsvang, har skrevet om biologisk kontroll av hhv. ugras, plantesjukdommer og skadedyr (kap.4). Dessuten har Atle Wibe (Bioforsk Økologisk) og Sverre Kobro (Bioforsk Plantehelse) skrevet hhv. tekstboks 4.12 og 4.13. Grete Lene Serikstad (Bioforsk Økologisk), Guro Brodal (Bioforsk Plantehelse), Trygve Rygg (pensjonist, tidligere Bioforsk Plantehelse), Haldor Fykse (pensjonist, tidligere UMB/Bioforsk Plantehelse), Ole-Christian Østreng (Debio) og Arne Oddvar Skjelvåg (UMB) har hatt større eller mindre deler av manuset til gjennomlesning og vi vil takke for hjelpen og viktige innspill.
Dette bokprosjektet er finansiert gjennom ekstern støtte fra Statens landbruksforvaltning («Veiledningstiltak for primærprodusenter») og Landbruks- og matdepartementet («Forskningsmidler over jordbruksavtalen»), men hadde ikke vært mulig å gjennomføre uten stor velvilje fra Bioforsk Plantehelse, som har bevilget mye gjennom bruk av grunnbevilgning. Stor takk til alle instanser som har muliggjort dette prosjektet.
I en oppjaget verden hvor lettvinne løsninger ofte står høyt i kurs, så vil kanskje enkelte mene at vi har gått vel mye i dybden på deler av denne boken. Vi håper allikevel at dere i ledige stunder har tålmodighet og nysgjerrighet til å gjøre dette «dypdykket» inn i materien rundt ugrasplanter, plantesjukdommer og skadedyr.
Ås, 22. januar 2006
Lars Olav Brandsæter
Dette kapitlet presenterer informasjon som gir en grunnleggende forståelse om skadegjørere, plantevern og plantehelse. Kapitlet beskriver bakgrunnen for økologisk landbruk og gir en kort gjennomgang av aktuelt regelverk og internasjonal organisering. Dette danner basis for å forstå hvordan økologisk landbruk i Norge er organisert og hvilke muligheter og begrensninger regelverket gir.
Økologisk landbruk bør betraktes mer som en utviklingsvei enn som en ferdig utviklet driftsmetode. Både regelverket og den agronomiske praksisen for denne driftsmetoden vil endres og utvikles videre etter hvert som kunnskap om økologi, biologi, agronomi og teknikk øker. Dette gjelder også ny kunnskap om produksjonsmetoders miljøkonsekvenser og kunnskap om samspill mellom maten vi spiser og vår helse.
Fra Debio har vi hentet følgende beskrivelse av idébakgrunn, overordna mål og praktiske forutsetninger mhp. økologisk landbruk:
"Økologisk produksjon bygger på et helhetssyn som omfatter de økologiske, økonomiske og sosiale sidene ved produksjonen, både i lokalt og globalt perspektiv. I det økologiske landbruket betraktes naturen som en helhet. I økologisk landbruk tilstrebes et selvbærende og vedvarende agroøkosystem i god balanse. Systemet baseres mest mulig på lokale og fornybare ressurser. Også ved videreforedling skal matvarene bearbeides ut fra det helhetlige utgangspunktet.
Grunnlaget for plantedyrkingen i hagebruk, jordbruk og skogbruk er hensynet til jordas struktur og fruktbarhet. I praksis betyr det blant annet å dyrke et mangfold av vekster og gjennomføre et variert vekstskifte der naturforholdene ligger til rette for det. Jordas fruktbarhet opprettholdes hovedsakelig ved resirkulering av gjødsel og annet organisk materiale.
Dyrkingssystemet utformes med flest mulig forebyggende tiltak mot skadegjørere. Lettløselig mineralgjødsel og kjemisk/syntetiske bekjempningsmidler brukes ikke. I økologisk landbruk tilstrebes balanse mellom dyretall og areal. Balanse betyr her at gården er mest mulig selvforsynt med fôr til husdyrene, og at tilførsel av gjødsel til jorda skjer med minst mulig risiko for forurensing av omgivelsene.
Ved foredling legges det vekt på å benytte skånsomme behandlingsmetoder, begrenset bruk av raffinering, energisparende teknologier og minimal bruk av tilsetningsstoffer. Genmodifiserte organismer, eller produkter framstilt ved hjelp av slike, er ikke tillatt i noen sammenheng".
Begrepet "økologisk landbruk" omfatter mer enn ordet økologi i vitenskapelig forstand. Økologisk landbruk, som er den norske parallellen til det engelskspråklige "organic farming", er en fellesbetegnelse for ulike driftsretninger. "Økologiforskriften" med veiledere setter dagens minimumskrav for hvordan driftsformen skal praktiseres. Regelverket for økologisk produksjon i Norge ble i utgangspunktet utviklet av Debio.
Gjennom EØS-avtalen har norske myndigheter ansvar for at Norge har en kontrollordning for økologisk produksjon. Herav følger det at norsk økologisk landbruksproduksjon må være i samsvar med EUs forordninger på dette området. Den første forordningen for økologisk landbruk kom i 1991. Etter den tid har det kommet ca. 40 forordninger på området som må bygges inn i det norske regelverket. EU-forordningene er forholdsvis detaljerte, og omhandler bl.a. regler for planteproduksjon, husdyrhold, kontroll og import.

I 1995 fastsatte Landbruksdepartementet for første gang forskrift for produksjon og merking av økologiske landbruksvarer. I forskriften ble Statens landbrukstilsyn og Statens næringsmiddeltilsyn (begge er nå en del av Mattilsynet) oppnevnt som kontrollmyndighet for primærproduksjon, pakking, foredling og import av økologiske landbruksvarer.
Debio er en medlemsorganisasjon som har utført kontrollvirksomheten på økologiske gårder siden 1986. I februar 1995 inngikk det som i dag er Mattilsynet en avtale med Debio, som går ut på at Debio har fått delegert myndighet for å utøve tilsyn med økologisk landbruksproduksjon i Norge. Dette innebærer bl.a. å følge standardrutiner ved kontroll og godkjenning av gårdsbruk og gartnerier. Debio inspiserer også foredlingsbedrifter på vegne av Mattilsynet. Begrepet økologisk landbruk er lovbeskyttet og varer kan bare omsettes som økologisk når produsenten har godkjenning fra Debio.
I svært mange land er økologisk landbruk på frammarsj, og selv om mangfoldet innen økologisk dyrking er enormt, finnes det faktisk et felles verdigrunnlag, uttrykt gjennom retningslinjer fra IFOAM (International Federation of Organic Agriculture Movements), den internasjonale sammenslutningen for økologisk landbruk. Blant annet er ønsket om å utvikle et bærekraftig landbruk, mest mulig basert på lokale og fornybare ressurser et felles, internasjonalt fundament. Et annet felles internasjonalt ståsted er å se landbruk og matproduksjon i et globalt og langsiktig perspektiv hvor føre var-prinsippet er en viktig grunnpilar.
Informasjon om regelverk som er relevant for økologisk planteproduksjon vil du finne mer om lenger bak i kapitelet.
Dyrking av jorda betyr store inngrep i det opprinnelige naturgrunnlaget. I økologisk landbruk søker man å opprettholde et balansert samspill mellom jord, mikroorganismer, planter, dyr og mennesker og på en slik måte opprettholde natur og miljø som grunnlag for landbruksdrift på lang sikt.
Landbruket blir stadig mer intensivt og spesialisert. Innkjøp av driftsmidler har økt kraftig, mens behovet for arbeidskraft har gått ned. Vi har fått et landbruk som produserer mer til lavere pris, men som kan ha flere negative konsekvenser, for eksempel:
Man kan si at økologisk landbruk har oppstått som en "motreaksjon" på denne utviklingen, og målsetningene for økologisk landbruk blir da også relatert til mange av de samme konsekvensene, men med "motsatt fortegn", dvs. at man ønsker å:
En vanlig oppfatning er at hovedforskjellen mellom økologisk og konvensjonelt drevet landbruk er at man ikke bruker plantevernmidler eller kunstgjødsel. Dette er imidlertid en svært forenklet måte å se det på. Mer vesentlig er at det legges vekt på alle faktorenes gjensidige avhengighet i driftsopplegget. Man skiller gjerne mellom økologiske, integrerte og konvensjonelle dyrkingssystemer.
Konvensjonell dyrking kjennetegnes ved utbredt bruk av innsatsfaktorer (for eksempel handelsgjødsel, plantevernmidler, kraftfôr) levert til gården utenfra, stor produksjon av bl.a. planter, melk og kjøtt som går ut fra gården, samt stor grad av mekanisering og ensidig drift. Det er som regel en driftsform med vekt på maksimering av gårdsinntekt med bruk av alle tillatte innsatsfaktorer.
Et økologisk dyrkingssystem er basert på et bedre definert verdigrunnlag. Dette synliggjøres bl.a. ved at:
I et integrert produksjonssystem tar en gjerne i bruk elementer fra både økologisk og konvensjonell drift ved at man f.eks. bruker plantevernmidler, men sprøyter etter behov og kombinerer med andre aktuelle tiltak (resistente sorter, mekaniske eller termiske bekjempelse etc.). Alle systemene er imidlertid under utvikling, og det finnes overganger mellom systemene.
På to svært sentrale områder, næringsforsyning og plantevern, skiller økologisk landbruk seg fra konvensjonell dyrking. Vedrørende næringsforsyning er det en målsetning for økologisk produksjon at eksport av næringsstoffer ut fra driftsenheten skal være minst mulig og at næringsstoffer til og fra planter og dyr går mest mulig gjennom en "lukket syklus". Dette kan blant annet oppnås ved hjelp av et gjennomtenkt vekstskifte, forsvarlig gjødselbehandling, og tilførsel av næringsstoff i samsvar med opptak i planter og varetransport ut fra gården.
Når det gjelder miljøproblemer knyttet til plantevernmidler, inklusiv rester i landbruksproduktene, er det lett å se miljøgevinsten ved økologisk produksjon: Syntetiske plantevernmidler brukes ikke i økologisk landbruk og driftsformen vil slik ha klare miljømessige fortrinn i forhold til konvensjonell produksjon. Det er imidlertid uenighet om hvor store konsekvenser restmengder av plantevernmidler i matvarer og i miljøet har. I økologisk landbruk kan bruken av plantevernmiddel med naturlig opphav, for eksempel planteekstrakter, være aktuelt, men bruken av disse kan også være omstridt.
Økologisk landbruk vil, som beskrevet over, i seg selv kunne ha flere fortrinn i forhold til konvensjonell produksjon, men forskning og praksis i økologisk landbruk kan også komme med løsninger som kan gjøre det konvensjonelle landbruket mer miljøvennlig, f.eks. bedre håndtering av husdyrgjødsel, optimal bruk av belgvekster og ugrasharving.
Før det moderne, kjemibaserte landbruket vokste fram, kan vi si at alt landbruk var økologisk drevet og stort sett erfaringsbasert med ingen eller begrenset kunnskap om grunnleggende prosesser. Var det ikke problemer med skadegjørere i landbruket på denne tida? Jo, Bibelen og andre historiske kilder tyder på at ulike skadegjørere var et kjent fenomen (blant annet i Am 4.9 og 5 Mos 28.22). Mennesker hadde altså kjennskap til sjukdommer og andre skadegjørere på planter før det moderne landbruket begynte å utvikle seg. Forklaringen ble lenge søkt i det overnaturlige. Også Romerne mente plantesjukdommer skyldtes guddommelige inngrep. Kongen Numa Pompilius (rundt 700 f.Kr.) innstiftet en årlig fest (Rubigalien) hvor en rød hund ble ofret for å avverge epidemier av rustsjukdommer i korn. Dette foregikk helt til begynnelsen av vår tidsregning. Det tok lang tid før folk flest ble bevisst at årsak til avlingssvikt skyldtes ulike skadegjørere.
I ulike historiske epoker har man søkt å løse plantevernproblemene med ulik tilnærming. Etter en periode med ervervelse av kunnskap om biologien til skadegjørerne tok man opp kampen på en mer rasjonell og vitenskapelig basert måte. Først ble kampen rettet direkte mot skadegjøreren med lite hensyn til uheldige bieffekter, men etter hvert ble tilnærmingen til fagområdet "plantevern" bredere, og i dag ser vi at fokus gjenspeiler hele spekteret fra enkeltløsninger til overordnete systemtilnærminger. Trenden i dag er å fokusere mer på systemløsninger, og her har økologiske landbrukere allerede lang erfaring. Plantevern inkluderer således alle tiltak som hindrer en skadegjører (ugras, insekter, midd, nematoder, pattedyr, virus, bakterier, sopp) i å redusere avlingen kvalitativt eller kvantitativt.
Av noen brukes i dag begrepet "plantehelse" ganske synonymt med begrepet plantevern, noe som kanskje ikke alltid er like velegnet. Vi kan kort definere at god plantehelse betyr planter som er sunne og friske (tekstboks 1.1).

I andre halvdel av 1800-tallet og fram til begynnelsen av 1900-tallet dreide fagområdet plantevern seg generelt om kunnskap om ulike skadegjørere som sjukdomsorganismer og skadedyr. Senere ble problemene med ulike skadegjørere mer og mer vinklet omkring begrepet plantevern. Dette viser en sterkere fokusering på å verne plantene mot skadegjørere ved hjelp av ulike tiltak. De seinere årene har begrepet plantehelse blitt mer vanlig. Dette gjenspeiles bl.a. ved at Plantesjukdomsloven (lov om tiltak mot plantesjukdommer og skadedyr på planter fra 1964) ble endret til Lov om plantehelse i år 2000. Det gir uttrykk for en økende forståelse for at målet ikke bare er planter uten synlig skade, men at plantene som helhet er sunne, dvs. at de har god motstandskraft mot skadegjørere. Fra 2004 ble Plantehelseloven innlemmet i Matloven.
Et problem med bruk av plantehelsebegrepet som en erstatning for plantevern, er at dette begrepet er mer relatert til sjukdomsorganismer enn til skadedyr og ugras. Spesielt i forhold til ugras føles begrepet plantehelse lite dekkende. Skadevirkningen av ugras er jo hovedsakelig avlingsnedgang på grunn av konkurranse og da passer ordet «plantehelse» dårlig. Det synes derfor mest naturlig å betrakte begrepet «plantehelse» som en del av fagområdet «plantevern».
Fagområdet plantevern er en form for «anvendt biologi». Biologisk kunnskap om kulturplantene og de ulike skadegjørere, og hvordan ulike kulturtiltak påvirker samspillet mellom kulturplante og skadegjører, er grunnlaget for utviklingen av fagområdet plantevern. Grunnleggende biologisk kunnskap er derfor avgjørende for alle som arbeider innen fagområdet plantevern, dette gjelder ikke bare forskeren, men også i høyeste grad bonden.
Selv om det i økologiske driftsformer legges vekt på å utvikle dyrkingssystemer mest mulig basert på naturens premisser, innebærer ikke det at en økologisk driftsform er et naturlig økosystem. Både den konvensjonelle og økologiske driftsform er i utgangspunktet menneskeskapt, med bl.a. matproduksjon for øyet. Et slikt kunstig økosystem, der mennesket, kulturplantene og det øvrige biotiske (planter/ugras, dyr/skadedyr, sopper, bakterier, virus) og abiotiske miljø (klima, næringsstoffer, vann, gasser) inngår, kalles et agroøkosystem (tekstboks 1.2). På grunn av forstyrrelser i et slikt system, som vekstskifte, pløying og andre agronomiske tiltak, har kulturmark en del fellestrekk med såkalte pionerplantesamfunn, der planter nyetablerer seg på åpen jord, for eksempel etter skogbrann, jordras etc. Å studere forekomst og utvikling av naturlige økosystemer gir økt forståelse for hvordan de ulike organismene fungerer i forhold til hverandre, også i et agroøkosystem. For naturlige økosystemer blir det lite relevant å snakke om skadeorganismer eller skadegjørere, siden man i stor grad kan si at de ulike organismene er avhengige av hverandre i slike systemer. Begrepet skadegjører (ugras, skadedyr og plantesjukdommer, eller uønskete organismer), er i agronomisk forstand bare relatert til kulturplantene, og det er derfor meningsløst å operere med dette begrepet i naturlige økosystemer.
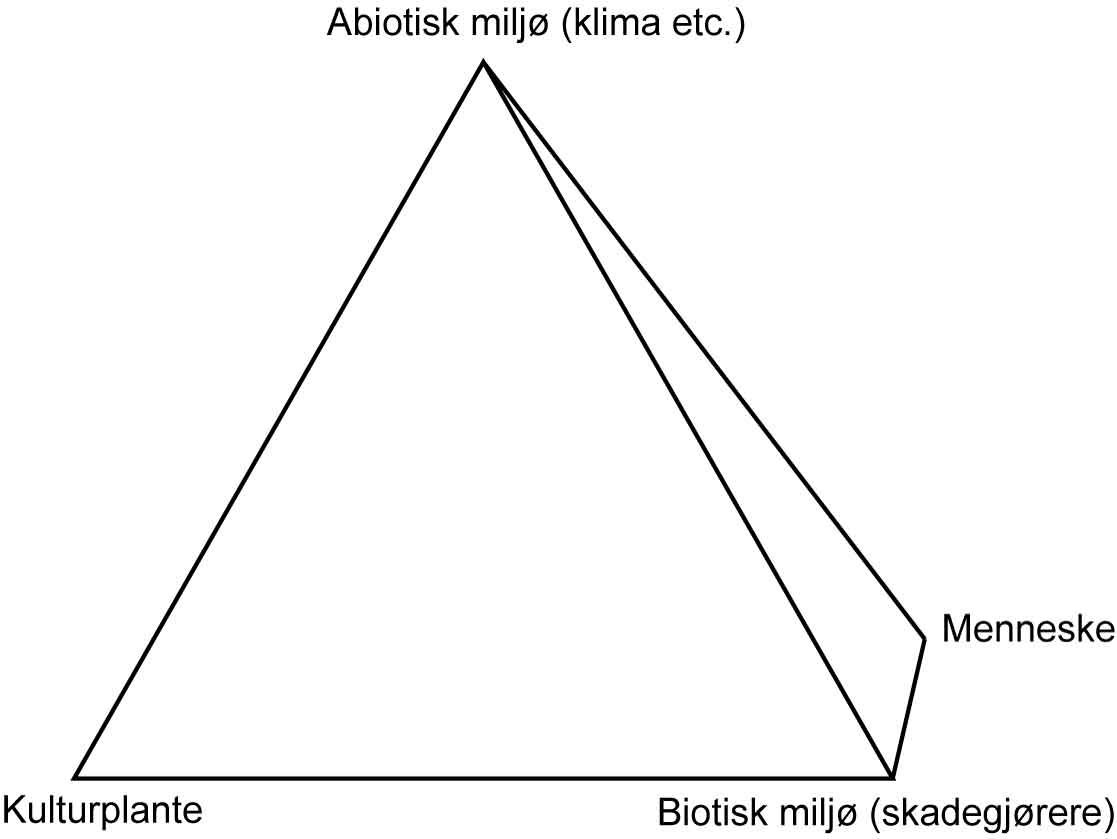
Figur 1.1 Et kunstig økosystem, der mennesket, kulturplantene og det øvrige biotiske (planter/ugras, dyr/skadedyr, sopper, bakterier, virus) og abiotiske miljø (klima, næringsstoffer, vann, gasser, jord) inngår, kalles et agroøkosystem. Disse faktorene og samspillet mellom disse blir ofte vist ved et tetraeder med de ulike faktorene («aktørene») i hvert sitt hjørne.
Innen fagområdet plantevern er det en evig søken etter å påvirke samspillet mellom kulturplantene, det abiotiske/biotiske miljøet og mennesket for å oppnå best mulig produksjon av kulturplanter i agroøkosystemet.
I tillegg til hjørnene/faktorene i tetraederet er det to elementer til: tid og rom. Fordi skadegjørerne utvikler seg over tid og ikke angriper alle planter eller arter samtidig, betyr det at skadegjørerne ikke finnes i samme grad overalt i én og samme kultur. Tilstedeværelse av skadegjørere betyr ikke alltid at det er behov for å sette inn tiltak. Skadegjørerne vil alltid være et naturlig element i et agroøkosystem. Det er betydningen de har for produksjonen som er avgjørende for om det er behov for sette inn tiltak. En slik vurdering krever
kunnskap om de enkelte skadegjørere, deres levevilkår, potensielle muligheter for å utvikle seg over tid (eventuelt over flere år) og i rom. Potensielle avlingsskader og økonomiske tap som angrep/konkurranse fra skadegjørerne kan medføre, må også taes med i vurderingen.
Om våren kan en oppdage brune prikker på undersiden av blad på hvitveis (figur 1.2) og gule flekker på vårkål. Dette er angrep av de respektive rustsoppene Tranzschelia anemones og Uromyces ficariae. Friske og angrepne planter kan stå ved siden av hverandre. Dette skyldes blant annet genetiske forskjeller i vertsplantepopulasjonen. Organismer som vi kaller «skadegjørere» er i utgangspunktet ledd i naturlige økosystemer. Skadegjørere som finnes i naturlige økosystemer, men som ikke er dominerende og iøyenfallende kan få en helt annen rolle gjennom menneskelig aktivitet, for eksempel gjennom en større genetisk likhet (mindre genetisk variasjon) i plantene som dyrkes. Dermed kan en naturlig skadegjører finne gode levevilkår på planter som er brukt til matproduksjon. Et godt eksempel er bringebærbladmidd som sjelden gjør mye av seg på villbringebær, men som kan gjøre stor skade på visse sorter av dyrket bringebær.
Nyoppståtte problemer med skadegjørere er ofte forårsaket av menneskelig aktivitet. Det betyr at bl.a. (potensielle) skadegjørere er blitt overført til økosystemer hvor de opprinnelig ikke var til stede. En del av disse skadegjørerne finner levevilkår i det nye økosystemet og kan utvikle seg til et plantevernproblem, mens andre ikke klarer seg i det nye økosystemet. På denne måten har mange planteskadegjørere spredt seg over hele verden. Man har som regel ikke vært klar over at et nytt problem kunne oppstå før skaden var et faktum. Risiko for denne typen situasjoner ligger bak føre var-holdningen til bruk av genmodifiserte organismer (GMO) innen økologisk landbruk, særlig i åpne økosystemer.

Skadegjørere i agroøkosystemer
Den vesentligste forskjellen mellom et naturlig økosystem og et agroøkosystem er den styrende/forstyrrende rollen mennesket har i systemet. Grunnlaget for dette er å forvalte areal, drive butikk og tjene til sitt levebrød.
Spørsmålet om hvorfor vi har skadegjørere i våre landbrukssystemer er på mange måter viktigere og mer interessant enn spørsmålet om hvordan vi kan bli kvitt skadegjørerne. Dette fordi svaret på det første ofte vil lede til et naturlig svar på det andre. Paradoksalt nok legger ofte mennesket/bonden forholdene til rette for at skadegjørerne kan etablere seg og utvikle seg videre. Det er disse menneskeskapte forholdene i samspill med skadegjørernes egenskaper, som gjør at problemet utvikler seg, og dermed bestemmer omfanget av problemet. Det er likevel viktig å innse at uansett hvor mye vi arbeider forebyggende mot skadegjørere, så vil en del skadeorganismer opptre i forskjellige kulturvekster.
Noen viktige egenskaper ved tradisjonelle/konvensjonelle agroøkosystemer, og eksempler på innvirkning på skadegjørere:
Kjenn din gård
Det er bonden selv som kjenner sin gård best. Hver gård har sin egenart, det kan for eksempel ligge jordstykker nær en elv eller bekk hvor dette skaper særegne klimatiske forhold. Andre deler av gården kan ligge høyere opp i lia eller ha et helt annet jordsmonn. Avstand til omkringliggende vegetasjon, for eksempel skogkanten, kan ha betydning. Tidligere erfaringer på gården, gjort over en rekke år, er viktig informasjon for hvordan gården kan utnyttes optimalt. Spesielt gjelder dette ved endringer i arealbruk eller driftsopplegg på gården.
Skade, avlingstap og økonomisk tap
Det økonomiske sluttresultatet for hele gårdsdriften er viktig for en gårdbruker. I tillegg vil også mange andre mål, som for eksempel trivsel med arbeidsoppgavene, være viktige. Det er også av stor betydning at en produsent ikke har et alt for ensidig syn på skadegjørere. Det er viktig å forstå forskjellen på begrepene skade, avlingstap og økonomisk tap.
En skade (eller symptom) på en plante kan skyldes biologiske (biotiske faktorer, hvor blant annet skadegjørere hører hjemme) og/eller ikke-biologiske faktorer (abiotiske faktorer, for eksempel klima og jordstruktur). Skader kan føre til avlingstap, dvs. redusert avlingsmengde (vekt eller volum) eller produktkvalitet. Det er imidlertid ikke alltid slik at skade gir avlingstap. For eksempel tåler agurk opptil 20 % angrep av mjøldogg på bladverket før det gir avlingstap. Avlingstapet kan føre til økonomisk tap.
Skade som oppstår ved lagring, transport og i detaljistleddet kan også være svært store. På verdensbasis blir denne formen for skade likestilt med skade som oppstår i selve produksjonsprosessen. Dette aspektet er vanligvis for lite belyst i debatten om verdens matvareproduksjon og matforsyning.
Sammenhengen mellom avlingstap og økonomisk tap er sammensatt. Økonomien henger sammen med markedet, og markedet reagerer også på avlingstap. Vanligvis reagerer et marked med økte priser når volumet av en vare reduseres. Hvordan dette igjen påvirker økonomien hos en enkeltprodusent, henger sammen med om denne produsenten har mer eller mindre skade enn andre produsenter. I et åpent marked kan reduksjon av produksjonsvolum bli fullstendig kompensert av høyere priser. Når skaden ytrer seg som kvalitetstap (kvalitetsforringelse) og ikke påvirker avlingsvolumet, kan det endelige økonomiske resultatet være svært lite tilfredsstillende når produkter av dårlig kvalitet erstatter høykvalitetsprodukter.
Samspill mellom skadegjørere og planter
En bør naturligvis prioritere å forhindre, eller i alle fall begrense, at skader oppstår. Et viktig emne er forståelse av samspill mellom planter og skadegjørere, og hvilke faktorer som påvirker dette samspillet.
De ulike skadegjøreres livsstrategier
For alle organismer kan vi gruppere deres livsstrategier i to deler:
Kunnskap om skadegjøreres livsstrategier er av stor betydning for utvikling av metoder for å regulere forekomst av slike organismer i landbruksproduksjonen.
Utvikling av skadegjørere
Alle utviklingsprosesser, i høy grad også populasjonsutviklingen til skadegjørere, har en viktig felles dimensjon, som er tid. Enhet for tid kan være sekund, minutt, time, dag, men også år og tiår. I plantevernsammenheng er time, dag og år ofte de mest relevante enhetene. I agronomisk sammenheng forholder man seg ofte til en vekstsesong, men på den annen side er det viktig å være klar over at for mange skadegjørere er det relevant å vurdere utviklingen over flere år. Oppbygging av jordboende smittekilder av ulike nematodearter eller frøbankens størrelse er gode eksempler.
Den andre viktige dimensjonen i utviklingen av skadegjørerer er utviklingen i rom, dvs. hvordan skadegjøreren sprer seg utover arealet. Eksempel her er i hvilken grad en "rose" (flekk) av åkertistel vokser i størrelse/diameter. Kombinerer vi de to dimensjonene, tid og rom, kan vi altså definere for eksempel hvor raskt en flekk av åkertistel vokser. I denne sammenhengen er det også viktig å nevne at en skadegjører aldri er jevnt fordelt over et område, og at denne fordelingen endrer seg over tid.
Grunnleggende kunnskap om utviklingen av skadegjørere finnes innen fagområdet populasjonsdynamikk (tekstboks 1.4).
Populasjonsdynamikk er en disiplin innen faget økologi. Populasjonsdynamikken har som mål å beskrive antallsendringer i forskjellige populasjoner mht. faktorene tid og rom. Metoder som brukes omfatter bl.a. observasjoner, kartlegging og kvantifisering av skadegjørerpopulasjoner. Innenfor populasjonsdynamikken studerer man også sammenhengen mellom populasjoner og dens «levende» og «døde» omgivelser. Innenfor fagområdet økologi kan vurderinger foregå på enkeltindivider (for eksempel et insekt eller en ugrasplante), populasjon (for eksempel alle kålfluene i en kålåker) og samfunnsnivå (man studerer flere arter samtidig). Gjennom analyser forsøker man å forklare hva som skjer. Ved å lage modeller kan man danne seg en tallmessig forståelse og dermed studere effekten av ulike inngrep mht. populasjonsutviklingen i samspill med omgivelsene. Kunnskap innen dette fagområdet betyr mye for forståelsen av hva som skjer med skadegjørere. Det danner grunnlaget for å iverksette tiltak for regulering av skadegjørere.
Populasjonsegenskaper
Populasjoner kan karakteriseres gjennom en del kvantitative og kvalitative egenskaper. I studiet av populasjoner ser vi at populasjonsegenskapene blir ganske forskjellig fra enkeltindividets. Hvert individ går gjennom en livssyklus, og de forskjellige livsfaser betyr mye for betydningen av hvert individ i utviklingsprosessen av hele populasjonen. For eksempel vil en voksen marihøne spise mindre bladlus enn sene larvestadier. Andel av sultne individer, altså sene larvestadier i en populasjon, er derfor avgjørende for den totale effekten. Det er viktig å være klar over hva som tas med i vurderingen, i dette tilfelle altså utviklingsstadiet til skadegjøreren, når en vurderer en hel populasjon.
Å kvantifisere populasjoner er ikke alltid enkelt. Individuelle ugrasplanter, insekter og nematoder kan på visse stadier tallfestes rimelig bra. Med sopp, virus og bakterier er det vanskeligere. For slike skadegjørere er det lettere og oftest mer vanlig å kvantifisere antall synlige symptomer, for eksempel antall bladflekker.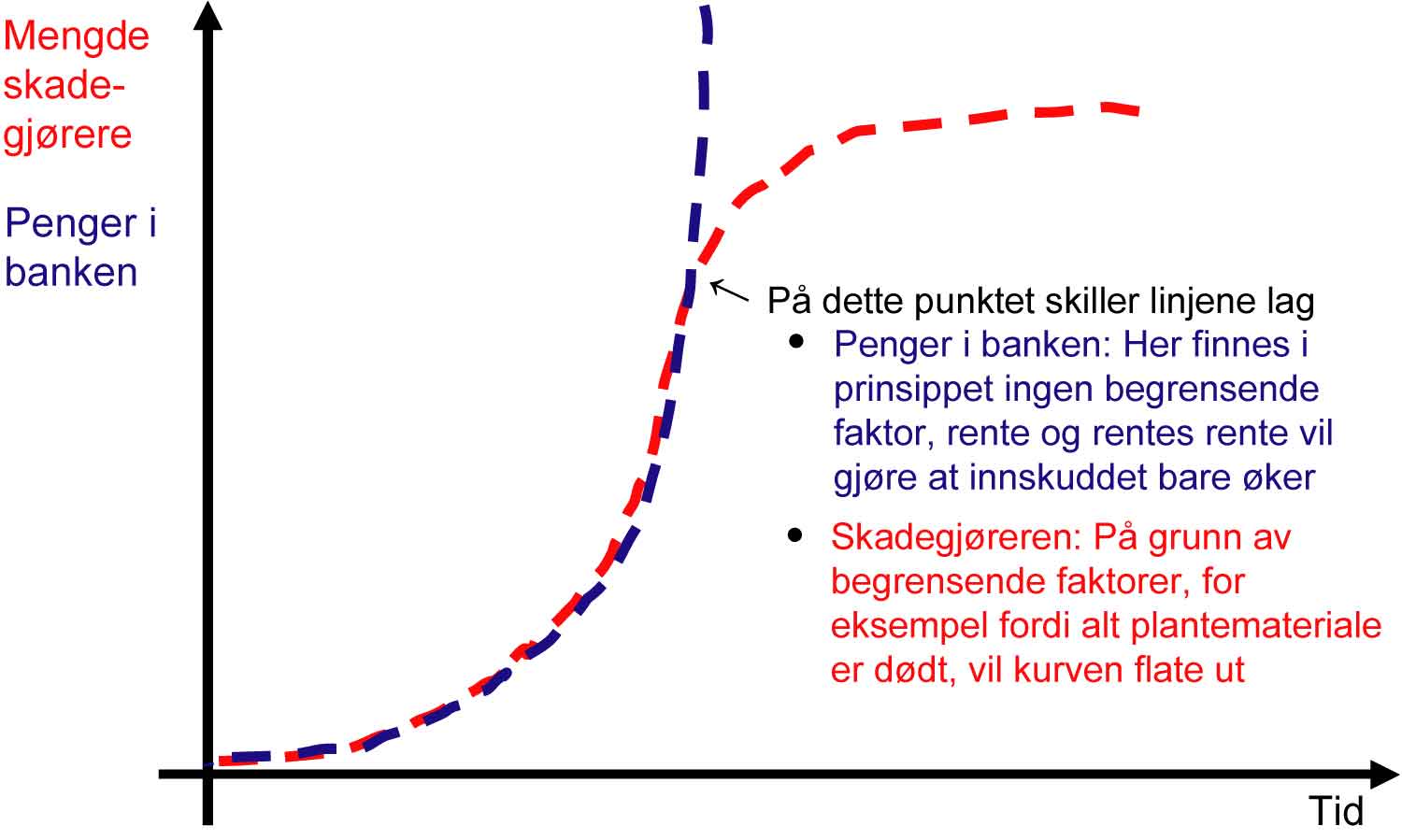
Generelle regler for populasjonsdynamikk
Som alle andre levende organismer har skadegjørere mulighet til å utvikle seg både ved at hvert enkelt individ vokser i størrelse, men også at antall individer i en populasjon øker. Denne prosessen kan sammenlignes med pengene som du har «på bok» i banken (se figur 1.3). Her vil hver krone gi en viss avkastning gjennom rentesatsen. Den endelige kapitalen vil avhenge av det opprinnelige innskuddet og rentesatsen. I tillegg vil det selvfølgelig være viktig om man underveis tar noe ut av banken.
Prinsipielt skjer det samme med skadegjørere som utvikler seg. Primærsmitten (utgangspopulasjonen), er her parallellen til innskuddet i banken, og utviklingsraten er tilsvarende rentesatsen som bestemmer hvor fort populasjonen vokser. Vekstprosessen i naturen fortsettes vanligvis ikke i det uendelige, men blir begrenset av en eller flere faktorer som gjør seg gjeldende etter hvert. At veksten tar slutt fordi temperaturen blir for lav, at det blir vanskeligere å finne friske plantedeler for skadegjøreren, og at modningsprosessen i planten tiltar, er eksempler på faktorer som påvirker utviklingsraten. Hvis vi skjematisk skal tegne en slik prosess med en graf, med tid på x-aksen, og angrepsnivå på y-aksen, får man i prinsippet en S-formet kurve. Utviklingsprosessen flater ut til et nivå som ofte blir betegnet som systemets bæreevne.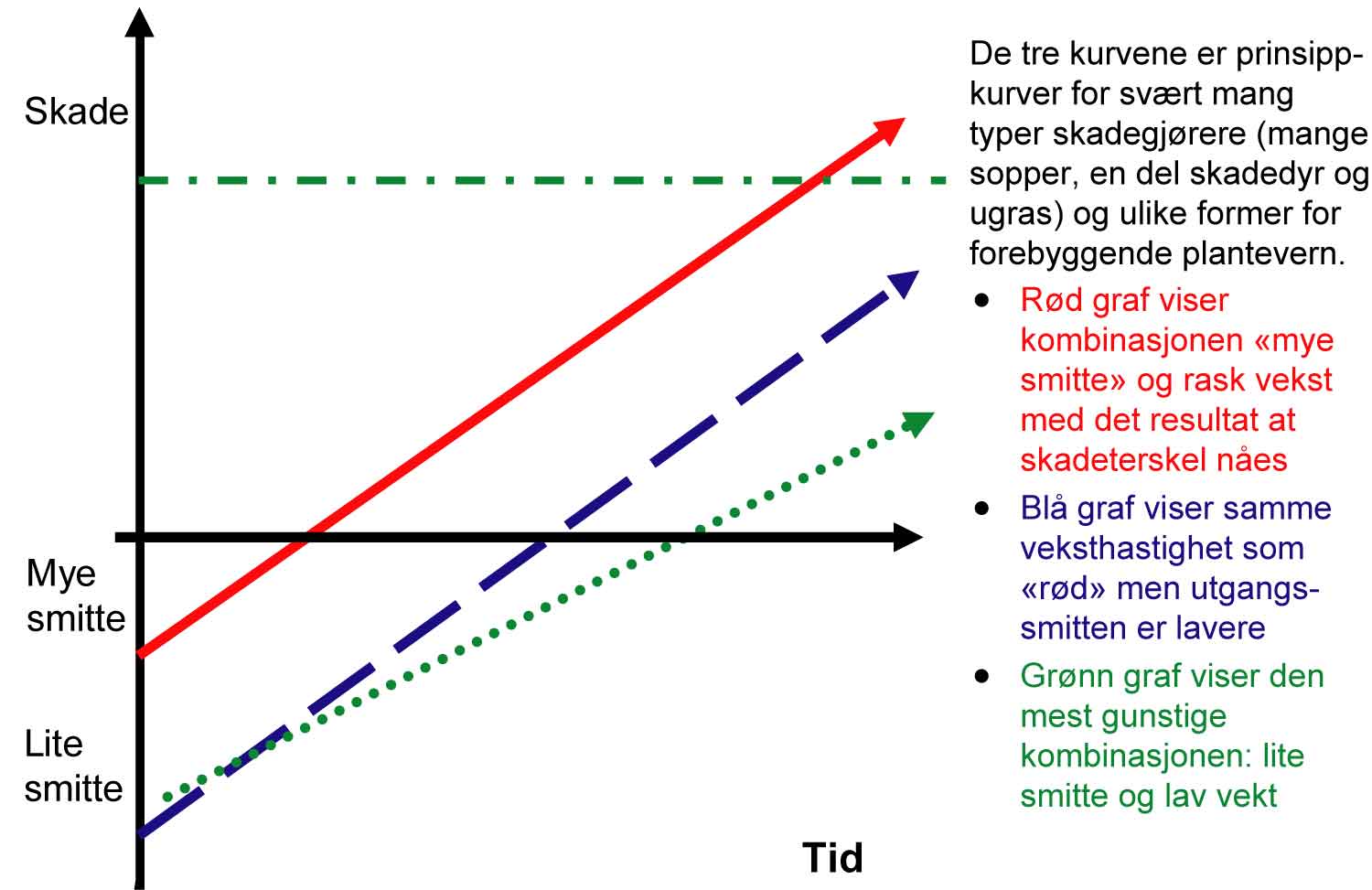
For lettere å forstå betydningen av primærnivået (det opprinnelige innskuddet i banken) og utviklingsraten (rentesatsen) i en utviklingsprosess av en skadegjører kan en gjengi den S-formede kurven gjennom en annen matematisk form på en slik måte at den kurvede linjen blir en rett linje. Ved å gjøre dette blir primærnivået og utviklingsraten straks synlig på en enkel måte. Primærnivået er nivået på vertikalaksen på tid null. Utviklingsraten gjenspeiles av hvor bratt den rette linjen blir over tid (rød graf på figur 1.4).
Nå kan vi «leke» med denne figuren. Hvis for eksempel primærsmitten blir lavere gjennom bruk av friskere plantemateriale, starter utviklingen på et lavere nivå og det tar lengre tid før det samme nivå (les skade) blir oppnådd (blå graf på figur 1.4). Hvis en samtidig endrer utviklingsraten gjennom bruk av for eksempel en mer resistent sort (jfr. lavere rentesats) går utviklingen tregere, altså en mindre vinkel mellom utviklingslinje og tidsaksen (grønn graf på figur 1.4). De ulike tiltak i en produksjon har mer eller mindre innvirkning på primærnivået av skadegjørere og/eller utviklingsrate av dem.
Populasjonsdynamikk og miljø
Ulike klimafaktorer som temperatur og fuktighet påvirker i stor grad etablering og utvikling av mange skadegjørere. Eksempelvis vil en periode med dugg på blader være avgjørende for spiring og inntrenging av noen typer soppsporer. For ugras er også miljøfaktoren lys viktig, for eksempel trenger frø av mange ugrasarter lys for å spire. Det kan være stor variasjon mellom ulike skadegjørere i hvordan de reagerer på samme miljøfaktor. For eksempel har ulike grupper av sjukdomsorganismer forskjellig pH-preferanser. Det samme gjelder temperatur og fuktighet, og det optimale kan variere for ulike arter.
Miljøet rundt en plante/skadegjører kan vurderes på tre ulike nivåer:
Klimaet i de 3 ulike miljønivåene vil være under stadig forandring, men vanligvis innenfor en viss og rimelig godt kjent variasjonsbredde. Endringer i miljøet kan gå raskt (sekunder/minutter) eller langsomt (opptil år). Det er viktig å ha en forståelse for hvilke faktorer i miljøet som er av betydning for skadegjørerens utvikling, og hvordan de gode miljøforholdene varierer.
Nøkkelfaktorer
Utviklingen av en skadegjører og omfanget av skaden den forårsaker, påvirkes ofte av flere faktorer. For de ulike kulturvekster og tilhørende skadegjørere er kunnskap om disse faktorene sentrale i arbeidet med å utforme effektive forebyggende tiltak og strategier. Å klarlegge slike nøkkelfaktorer, som kan hjelpe oss i kontrollen av skadegjørerne, er derfor et viktig arbeid.
Skadegjørere, det være seg sjukdomsorganismer eller skadedyr, har alle sin plass i næringskjeden. Litt forenklet kan vi si at dette betyr at alle spiser og blir spist. Med litt fantasi kan man kanskje også integrere ugraset i slike næringskjeder, men da må vi oppfatte ordet "spising" som et utvidet begrep. De ulike næringskjedene kan være veldig komplekse og dynamiske, særlig når vi vurderer antall og antallsendringer over tid innen hvert enkelt ledd (for eksempel en skadegjører) av næringskjeden. Det er viktig å utvikle en forståelse for hvordan slike endringer i næringskjeden oppstår og hvordan disse kan påvirkes. I denne sammenhengen er det viktig å huske at det verken er mulig eller ønskelig å endre naturen gjennom å forandre naturens lover, men at våre påvirkningsmuligheter består i til en viss grad å kunne endre de faktorene som styrer omfang og hastighet av prosesser i naturen. Slike menneskestyrte endringer kan medføre både nytte og skade i landbrukssystemer, og det er viktig å være klar over at langsiktige prosesser er vanskelige å bedømme på forhånd.
De styrende faktorene finnes både i den biotiske (levende) og abiotiske ( ikke-levende) natur. De biotiske faktorer kan grupperes i to:
De fleste planter er «immune» for de fleste skadegjørerer. Likevel er det mange mikroorganismer, insekter, midd og nematoder som kan skade planter. Innenfor en enkelt planteart (for eksempel bygg) er det sorter med forskjellig motstandsevne og det finnes forskjellige genetiske varianter (raser) av skadegjørerene. Sortsvariasjon i kulturplantene og rasevariasjon i skadegjørere vil følgelig gi variasjoner i angrepsgrad. Samspillet mellom plante og skadegjører betegnes med forskjellige begreper:
Aggressivitet hos skadegjører:
Aggressivitet betegnes som parasittens evne til å skade en vertorganisme.
Virulens hos skadegjører:
Virulens betegner skadegjørerens evne til å angripe en vertorganisme.
Patogenitet hos skadegjører:
En organisme, vanligvis brukt om sopp, bakterier og virus, sin evne til å forårsake sykdom. Patogeniteten til en organisme blir bestemt av både aggressivitet og virulens.
Resistens (=Motstandsdyktighet) i planter:
Resistens hos planter er evnen til å hindre skadegjørerens vekst og utvikling, dvs. redusere skadegjørerens evne til «å fungere som en skadegjører».
Mottakelighet (motsatt av resistens) hos planter:
En plante med høy mottakelighet har svak eller manglende evne til å begrense skadegjørerens vekst og aktivitet. Her snakker vi med andre ord om summen av egenskaper som tilrettelegger for at en organisme blir en vert for en parasitt.
I tillegg til egenskaper hos vert og skadegjører, er miljøfaktorer som temperatur, vann og næring avgjørende for omfanget av skaden som kan utvikles.
Biodiversitet
Begrepet biodiversitet blir stadig hyppigere brukt i forbindelse med landbruksproduksjon. Biodiversitet brukes for å karakterisere biologisk mangfold i et økosystem, dvs. forekomst av mange arter. Det legges ofte et verdigrunnlag i dette begrepet. Generelt vil man oftest forbinde en høy biodiversitet med noe positivt. Om dette alltid er positivt i landbrukssammenheng kan likevel diskuteres. Denne vurderingen, om det er positivt eller ikke, henger sammen med rollen biodiversitet spiller i et agroøkosystem. Begrepet funksjonell biodiversitet gir et visst uttrykk for denne presiseringen.
Agroøkosystemer med høy grad av biodiversitet blir ofte karakterisert som mer robuste enn systemer med lavere biodiversitet, ut fra den erkjennelse at nivået av skadegjørere blir holdt i sjakk av ulike styrende faktorer i systemet. I den grad høy biodiversitet er med på å løse plantevernproblemene vil det altså ofte være slike styrende faktorer som vi allerede har diskutert som er forklaringen bak dette.
Les mer i tekstboks 1.6.
Biodiversitet har betydning innenfor plantevern. Det henger sammen med vertsplantespekter og graden av spesialisering hos skadegjøreren. For eksempel storknolla råtesopp (Sclerotinia sclerotiorum) har et vertsspekter med flere hundre plantearter og er dermed ikke så kresen. Vekstskifte kan ikke så lett løse problemene med angrep av denne soppen hvis man ikke vurderer vertsplantespekteret nøye. Andre skadegjørere er spesialiserte, som for eksempel mange rustsopper, insekter og nematoder. Her har vekstskifte mye mer å tilby. Potetcystenematode angriper bare potet og nære slektninger av potet. Enkelte sopper kan være så spesialiserte at de bare angriper en sort og ikke andre innen samme planteart. Dette gjelder ofte genspesifikk resistens. Dette betyr ikke at økt biodiversitet på sortsnivå (altså mange forskjellige sorter) samtidig betyr stor diversitet av enkelte egenskaper. Her bruker man begrepet intraspesifikk biodiversitet. Når en snakker om biodiversitet som omtaler variasjon i arter snakker en om interspesifikk biodiversitet. Mange forskjellige sorter kan være nære slektninger og dermed bære nøyaktig de samme genene en bonde er interessert i.
Biodiversitet i tid og rom
Biodiversitet er ikke et statisk fenomen. Levende organismer, særlig i landbrukssammenheng, er i stadig endring. Dette varierer fra sted til sted. Erfaringer fra egen gård gir et visst inntrykk av slike variasjoner.
Biodiversitet og habitat- ( leveområde-) diversitet
Variasjon mht forutsetningene i leveområder er det viktigste grunnlaget for variasjon i levende organismer. Disse områdene kan endre egenskaper over tid. Disse endringene skjer ved faste naturlige rytmer, ( dag-natt, årssesonger) men også gjennom naturlige og menneskelig inngrep. Dette innebærer at biodiversitet til en viss grad kan styres, bevisst eller ubevisst i en riktig eller feil retning.
Målemetoder
Det finnes forskjellige måter å uttrykke biodiversitet på, men vanligvis brukes diversitetsindeks (DI). Det finnes flere utgaver også av denne, men alle er funksjoner av antall arter som er representert og antall
individer av hver art. Det er viktig å kjenne hva slags biodiversitet indeksen gjenspeiler for en riktig vurdering.
Funksjonell biodiversitet
En del av de viktige funksjonene som biodiversitet har i naturlige systemer, er blitt redusert i det moderne landbruket. Genetisk uniformt plantemateriale skaper ideelle forhold for utvikling av skadegjørere. Ny kunnskap forsøker å gjenvinne det tapte terrenget. Tiltak som øker biodiversiteten med det mål å utnytte de positive egenskapene, er bl.a. samplanting, underkulturer og sortsblandinger. Vekstskifte kan vurderes som et tiltak for økt biodiversitet, men her ligger biodiversitetens rolle på et lengre tidsperspektiv.
Et annet tiltak i strøk med stor andel av dyrket mark er å skape særmiljøer hvor naturlige fiender av skadegjørere trives.
Inntil slutten av 1800-tallet blandet offentlige myndigheter seg lite inn i styre og stell i landbruket. Etter hvert viste det seg at ugras, skadedyr og sjukdomsorganismer reduserte avlingene betraktelig, og et stort behov for både ny kunnskap gjennom forskning og reguleringer gjennom lovverk og kontroll, meldte seg. I kjølvannet av disse nye behovene ble W.B. Schøyen i 1891 ansatt som offentlig landbruksentomolog. I 1894 ble tittelen omgjort til statsentomolog med ansvar for både plantesjukdommer og skadedyr. Skrifter fra Schøyen til Landbruksdepartementet er meget verdifulle kilder til kunnskap om plantehelsen i Norge for disse årene. Den offentlige forvaltningen omkring planteskadegjørerne vokste: I 1913 ble således Emil Korsmo tilsatt som statskonsulent i ugrasspørsmål, og i 1919 ble en stilling som statsmykolog opprettet. I 1907 ble for første gang en planteskadegjører, stikkelsbærdreper, regulert gjennom offentlig lovverk. Dette ble betydelig utvidet i 1916 gjennom, Lov om bekjempelse av skadeinsekter og plantesjukdommer fra 1916. Det er en lang og komplisert historie fra denne pionértida og fram dagens forvaltning og lovregler, som i tillegg til skadegjørere også omfatter omsetning og bruk av plantevernmidler, inkludert økologiske.
Regelverk
Alt regelverk og all forvaltning omkring matproduksjon og mattrygghet, inkludert økologisk produksjon, er fra 2004 hjemlet i Matloven. Foruten å sikre helsemessig trygge næringsmidler og fremme helse, kvalitet og forbrukerhensyn langs hele næringskjeden, samt ivareta miljøvennlig produksjon, skal loven fremme god plante- og dyrehelse. Dette er en fullmaktslov som forvaltes av helse og omsorgsdepartementet (HOD). Mattilsynet fører tilsyn og fatter vedtak for gjennomføring av bestemmelsene i loven og tilhørende forskrifter. Spesielle krav og forpliktelser innen plantehelse er omtalt i matlovens §18. For å sikre gjennomføring og utfylling av loven er det fastsatt en rekke forskrifter. De viktigste på området planteproduksjon, plantevern/plantehelse og økologisk produksjon er kort omtalt nedenfor.
Forskrift om økologisk produksjon og merking av økologiske landbruksprodukter og næringsmidler fra 2005 skal sikre at landbruksprodukter og næringsmidler, som markedsføres som økologiske, overholder produksjonskravene. Videre skal forskriften ivareta miljøvennlig produksjon og sikre redelig omsetning. Mattilsynet har med faglig bistand fra Debio utarbeidet tre veiledere til forskriften:
Mattilsynet har tilsynsansvar for økologisk produksjon, foredling og salg av økologisk mat. Mattilsynet har imidlertid inngått avtale med Debio om gjennomføring av selve den praktiske kontrollen ute på gårdsbruk og i bedrifter. Debio rapporterer til Mattilsynet om resultatene av kontrollen.
Forskrift om planter og tiltak mot planteskadegjørere (01.12.2000, vil bli revidert i 2006). Formålet med forskriften er å hindre introduksjon og spredning av planteskadegjørere, bekjempe eller utrydde eventuelle utbrudd i Norge og sikre produksjon og omsetning av planter og formeringsmateriale med best mulig helse og tilfredsstillende kvalitet. Forskriften inneholder bestemmelser om tiltak mot planteskadegjørere, spesielle bestemmelser om produksjon og omsetning av planter og formeringsmateriale (registreringsordning, krav til kvalitet, identitet/ renhet, sertifisert produksjon), spesielle bestemmelser ved innførsel/utførsel (sunnhetssertifikat), avgifter osv. Forskriften har en rekke vedlegg, bl.a. lister over planteskadegjørere som det er forbudt å introdusere/spre i Norge, såkalte karanteneskadegjørere eller farlige skadegjørere. Ethvert funn av disse må straks meldes til Mattilsynet, og ved påvisning av slike gir Mattilsynet pålegg om gjennomføring av tiltak som skal hindre videre spredning og om mulig bekjempe skadegjøreren.
Bioforsk Plantehelse yter viktig faglig forvaltningstøtte, inkludert diagnose av en rekke planteskadegjørere og faglig rådgivning, til Mattilsynet på plantehelseområdet.
Det norske plantehelseregelverket er basert på at Norge har inngått flere internasjonale avtaler. Dette gjelder bl.a. den internasjonale plantehelseavtalen (International Plant Protection Convention, IPPC) som administreres av FAO og avtalen om veterinære og plantesanitære tiltak (Sanitary and Phytosanitary measures, SPS-avtalen) under verdens handelsorganisasjon (WTO). Sammen med Landbruks- og matdepartementet (LMD) deltar Mattilsynet i IPPC. Blant de viktigste oppgavene her er utvikling av internasjonale standarder for plantehelse.
På europeisk nivå deltar Mattilsynet, sammen med Bioforsk Plantehelse, i samarbeidet i den europeiske plantevernorganisasjonen European and Mediterranean Plant Protection Organisation (EPPO). Organisasjonen gir bl.a. anbefalinger om hvilke offentlige tiltak mot planteskadegjørere som medlemslandene bør sette i verk, samt anbefalinger om hvilke spesifikke krav som bør stilles ved handel med planter og plantedeler. Mattilsynet samarbeider også jevnlig med planteinspeksjonstjenestene i de andre nordiske landene, bl.a. gjennom årlige møter.
Plantehelse er per i dag ikke en del av EØS-avtalen. Det ser ikke ut til å være nødvendig så lenge aktiviteten i IPPC og EPPO dekker de viktigste problemstillingene.
Forskrift om såvarer fra 1999, og forskrift om settepoteter fra 1996, med seinere endringer, har som formål å sikre produksjon og omsetning av såvare og settepoteter med best mulig helse og kvalitet. Forskriftene m/vedlegg inneholder bl.a. betingelser for dyrking og produksjon og krav til kvalitet. Dersom såvaren/ settepotetene tilfredsstiller kravene ved vekstkontroll, kontrolldyrking og laboratorieanalyser blir såvaren/settepotetene sertifisert av Mattilsynet. Produksjon, innførsel og omsetning av såvare/settepoteter kan bare skje dersom betingelsene i forskriftene er oppfylt.
I henhold til veileder B i økologiforskriften skal økologisk godkjent såvare og formeringsmateriale benyttes. De samme kvalitetskravene gjelder for økologisk som for konvensjonelt dyrka såvare og formeringsmateriale. For ettårige vekster gjelder at produksjonen skal skje på sertifisert/godkjent økologisk areal (såing og høsting). For flerårige gjelder det at produksjonen skal være i henhold til driftsreglene i minst to år og at høsting skjer på godkjent økologisk areal (dvs. såing kan skje på karensareal). Dersom økologisk formeringsmateriale av ønsket sort ikke kan skaffes, kan ubeiset konvensjonell vare brukes.
Retningslinjer for sertifisert produksjon av hagebruksvekster fra 2001, er en frivillig ordning for å produsere friske planter av god kvalitet og riktig sort. Pr. 2005 omfatter retningslinjene bærvekster, prydplanter i veksthus og frukttrær og grunnstammer til frukttrær. Retningslinjene stiller strengere krav til både produksjonssted, produksjonsmetoder, dyrkingsmedier og hygiene, samt krav om fravær av flere skadegjørere, enn det plantehelseforskriften har som minimum. Bare virksomheter godkjent av Mattilsynet kan produsere sertifisert plantemateriale. Sertifiserte plantefelt blir regelmessig testet og kontrollert av Mattilsynet etter fastsatte retningslinjer.
Forskrift om plantevernmidler fra 2004 omfatter foruten de kjemiske plantevernmidlene også andre stoffer og biologiske preparater eller organismer som skal brukes til å verne, hemme eller forebygge angrep av planteskadegjørere. Også stoff eller preparat som blir utgitt for å regulere plantevekst (unntatt gjødselstoffer), drepe eller hemme uønska vekst av blader og ris, kontrollere uønska vegetasjon utenom kulturjord og øke virkningen av andre plantevernmidler, omfattes av forskriften. Plantevernmidler skal godkjennes av Mattilsynet.
Agronomisk utprøving av plantevernmidler utføres av Bioforsk Plantehelse på oppdrag fra Mattilsynet.
Floghavre er et ugras som er vurdert som så farlig for norsk korndyrking at det er underlagt egen forskrift, Forskrift om floghavre fra 1988. Dersom ugraset påvises, skal det bekjempes. Det er forbudt å omsette såvare, halm, helt korn og erter til fôr, husdyrgjødsel, kompost, jord mm fra eiendommer hvor det er påvist floghavre, dersom ikke varen er behandlet slik at spireevnen til eventuelle floghavrefrø er ødelagt. Mattilsynet fører et register over alle gårder der ugraset er påvist.
Forskrift om prøving og godkjenning av plantesorter fra 1999, fastsetter vilkår og fremgangsmåte for opptak av plantesorter på offisiell norsk sortsliste (utgis av Mattilsynet), og skal bidra til at sorter som markedsføres er best mulig tilpasset norske forhold. En plantesort kan bare oppføres på offisiell norsk sortsliste dersom den har bestått nyhetsprøving og gjennomført verdiprøving slik at den anses å ha en tilfredsstillende dyrkings- og bruksverdi i Norge. Verdiprøving av sorter utføres av Bioforsk på oppdrag fra Mattilsynet.
Oppdaterte lover og forskrifter med vedlegg finnes tilgjengelig på www.lovdata.no
Mattilsynets Nasjonalt senter for planter og vegetabilsk mat, ved regionkontoret for Oslo, Akershus og Østfold i Ås, koordinerer arbeidet med å opprettholde god plantehelse i Norge og redusere risikoen for introduksjon og spredning av planteskadegjørere med planter, plantedeler, trelast, jord m.m. og at innsatsvarene i planteproduksjon: frø og plantemateriale, plantevernmidler, gjødsel, kalk, dyrkingsmedier og kompost, har høy kvalitet og nytteverdi.
Kimen Såvarelaboratoriet AS (tidligere Såvarelaboratoriet i Landbrukstilsynet/ Statens frøkontroll) er autorisert av Mattilsynet til å utføre offisielle laboratorieanalyser av såvarer som grunnlag for sertifisering. I tillegg utfører laboratoriet analyser av såvarer på oppdrag fra såvarebransjen og dyrkere/bønder som bruker egen såvare. Laboratoriet er også autorisert til å diagnostisere floghavre og analysere for innhold av floghavre i helt frø til fôr.
Mer informasjon omkring tilsyn med planter, plantevern/plantehelse og økologisk produksjon finnes på www.mattilsynet.no og www.debio.no.
Regelverk for plantevern i økologisk produksjon
Regelverket omkring plantevern i økologisk landbruk er bygd opp rundt erkjennelsen av at det er viktig med forebyggende tiltak. Selv om det ikke er noen bestemte krav til sunnhetstilstand utover det som kreves for konvensjonell såvare og formeringsmateriale, er det viktig med så friskt og ugrasfritt utgangsmateriale som mulig ved økologisk produksjon.
Økologiforskriftens veileder B omhandler tiltak som skal anvendes for å forebygge og unngå ugras, skadedyr og plantesjukdommer:
Som vedlegg til veileder B finnes bl.a. lister over nytteorganismer som er godkjent til biologisk bekjempelse i økologisk produksjon (liste 2 B1) og liste over preparater som er tillatt brukt i økologisk produksjon (liste 2 B2). For noen av disse kreves forhåndsgodkjenning av Debio. Midlene og nytteorganismene i B1 og B2 er godkjent av Mattilsynet i henhold til Forskrift om plantevernmidler. Oppdaterte lister finnes på Mattilsynets internettsider ( www.mattilsynet.no)
Liste 2 C inneholder anbefalinger ved bruk av hjemmelagde planteuttrekk og preparater/plantestyrkende midler.
Som vi har vært inne på tidligere, vil risiko for skade i en produksjon være svært avhengig av hvilken produksjon det er snakk om, men også hvor produksjonen foregår. For eksempel vil produksjon av økologisk potet, pga. tørråtefaren, være langt mer risikofylt i nedbørsrike lavlandsstrøk i Sør-Norge enn mer tørre eller kaldere strøk. Økologisk produksjon av frukt og bær er også eksempler på kulturer hvor vi kan fastslå at risikoen, i alle fall i dag, er ganske stor. I motsetning til potet er det for eksempel for eple vanskelig å finne områder hvor produksjonen er lite utsatt for skadegjørere.
I det økologiske miljøet verserte det tidligere en del kontroversielle påstander omkring plantevern i økologisk landbruk, bl.a. at økologiske produkter var lite utsatte for skadedyr (se NO FILENAME).
Hva gjør bonden hvis et problem verken kan løses ved hjelp av forebyggende eller direkte tiltak? Den ytterste konsekvens kan være at en er nødt å vurdere om produksjonen passer for gården. Denne egnethet for en produksjon på en bestemt økologisk gård er et grunnleggende tema i planlegging av økologisk produksjon. På tysk har man et velegnet og mye brukt begrep for dette: Standortgerechter Anbau = Stedstilpasset produksjon. Potetproduksjon kan for eksempel være et problem der tørråte vanligvis forårsaker en tidlig nedvisning av potetriset. Tørråtesoppen fører ikke bare til avlingstap, men også til at næringsstoffene i jorda ikke blir utnyttet og derfor lett kan vaskes bort. Skrantende kulturplanter vil også øke ugrasproblemene. På enkelte gårdsbruk/ lokaliteter kan derfor konklusjonen bli at bonden bør kutte ut en produksjon helt og heller satse på andre kulturer.
Et annet aspekt omkring risiko handler om hvor avhengig bonden er av en bestemt produksjon. Hvis drifta på en gård i stor grad er bygd opp rundt en eller noen få store produksjoner, sier det seg selv at man er sårbar hvis en skadegjører med stor skaderisiko dukker opp og man ikke har aktuelle tiltak til rådighet. Et mer allsidig driftsopplegg vil derfor være en strategi mot at stor skade i en kultur skal ødelegge økonomien et gitt år. Dette er spesielt viktig i risikofylte produksjoner, for eksempel ved økologisk fruktproduksjon.
Finnes det løsninger for alle plantevernproblemer? Svaret på dette spørsmålet er sterkt relatert til hvor langt vi kan og er villige til å strekke oss for å gjøre driftsopplegget mest mulig robust. For eksempel vil et driftsopplegg med relativt ensidig kornproduksjon kunne gi grunnlag for flere plantevernproblemer, som flerårig ugrasarter som kveke og åkertistel. Et driftsopplegg med mindre korn, hvor eng og rotvekster inngår, vil være langt mer robust mht. flerårige ugrasarter. I driftsopplegg med mye korn vil dessuten kornsjukdommer som overvintrer på halmrester og en del jordboende skadegjørere kunne redusere stabiliteten. I et agronomisk perspektiv finnes altså tiltak/strategier for i alle fall å løse en del av disse problemene. Spørsmålet kan være om produsentene har mulighet eller vilje til å gjøre et slikt valg, og dette henger også sammen med prioriteringer som ligger utenom de agronomiske, for eksempel myndighetenes landbrukspolitikk, som er avgjørende for bondens økonomi.
I en del produksjoner vil det kunne finnes tiltak som minsker risikoen til det minimale, men som samtidig gjør produksjonen langt mer ressurskrevende både mht. driftsmidler og arbeidsbehov. Et eksempel på dette er bruk av fiberduk eller insektnett for å hindre insektangrep i flere utsatte grønnsakkulturer. Det avgjørende er om innsatsen blir godtgjort gjennom en reduksjon av forventet skadenivå.
Som en oppsummering av ulike faktorer som er viktige for utvikling av plantevernstrategier for å minimalisere risiko i økologisk landbruk, kan vi sette opp følgende hovedlinjer:
Et problem kan ofte være at teknikker som fungerer i mindre målestokk, for eksempel i en kjøkkenhage, ikke er gjennomførbare for større arealer. Driftsopplegg har mye å si for bondens plantevernsituasjon, men selv under optimale forhold vil bonden likevel ha mange utfordringer mht. å løse sine plantevernproblemer. Når det gjelder for eksempel ugras, vet man at uansett hvor optimalt driftsopplegg man har lagt opp til, vil man få total avlingssvikt i mange kulturer hvis man ikke setter inn direkte tiltak som fast rutine i driftsplanen. Et annet aspekt er at i mange kulturer finnes et mangfold av skadegjørere som potensielt kan utvikle seg, og de ulike skadegjørere kan reagere veldig forskjellig på ett og samme kulturtiltak. Det er med andre ord en dårlig løsning om man gjennom et bestemt tiltak løser et problem men skaper et annet. Som vi skal komme tilbake til senere, er det ikke bare agronomiske tiltak som påvirker skadegjørernes utvikling.
Fra å bekjempe til å forebygge...
Det å forebygge, eller hindre, at skadegjøreren dukker opp eller gjør skade, istedenfor å bekjempe skadegjøreren etter at den har dukket opp, er i tråd med ved selve grunntanken i økologisk landbruk. Det er derfor svært viktig å utvikle robuste strategier som bygger på mest mulig forebyggende plantevern. Men når dette er sagt, må det nok en gang poengteres at det også i mange situasjoner innen økologisk dyrking vil være helt nødvendig med direkte tiltak fordi det er umulig å forebygge alle problemer. Frøugras og andre skadegjørere med stor utviklings- og spredningsevne, for eksempel kålmøll, (ofte kombinert med liten motstandsdyktighet hos kulturveksten), er eksempler på dette.
Innen plantevern kan det skilles mellom tiltak som har kortvarig eller lengre virkningsgrad. Langsiktige løsninger får man ofte gjennom, la oss kalle det "strategiske" valg. Tidsperspektivet er da gjerne flere vekstsesonger. Et typisk eksempel på et slikt strategisk valg er planlegging av vekstskifte. Omløpsplaner må være satt sammen slik at de forebygger problemer med skadegjørere på lengre sikt. Mer kortsiktige valg kan vi kalle "taktiske" valg, og disse har i prinsippet bare et perspektiv begrenset til en vekstsesong. Valg av tidspunkt for radrensing er et eksempel på dette. Et annet taktisk valg kan gjelde sortsvalg. I en del tilfeller kan strategiske og taktiske valg være svært sammenflettet. Som eksempel kan nevnes: 1) Man hører ofte at det er viktig å ta med rotvekster i et vekstskifte for å kunne hanskes med de flerårige ugrasartene. Dette gjør man ikke fordi rotvekstene er spesielt konkurransesterke overfor for eksempel åkertistel. Årsaken er snarere at en radkultur gir gode muligheter for radrensing. Det mest effektive for kontroll av flerårige ugrasarter vil selvfølgelig være å bruke en konkurransesterk radkultur (eksempel: hodekål vs. gulrot/løk). Det strategiske og det taktiske valget kan henge nært sammen. En serie med effektive direkte tiltak mot flerårige problemugras samlet sett er et forebyggende tiltak hvis dette bidrar til å redusere problemet på lengre sikt. 2) Mht. jordboende skadeorganismer kan effekten av valg av resistent sort, for eksempel potetcystenematode-resistent potetsort, ha et langsiktig perspektiv i tillegg til den kortvarige. Sortsvalget det ene året gjør at smittenivået i senere år forhåpentligvis er under skadeterskel. Den endelige effekten er avhengig av vertsplantespekter og virulensegenskaper av de enkelte skadegjørere og dens samspill med den sorten som er valgt. Sortsvalg kan altså ha både en strategisk og en taktisk dimensjon. For bonden vil effektive forebyggende tiltak med langvarig virkning ofte være økonomisk gunstige. Generelt kan vi si at desto mer en plantevernstrategi er basert på forebyggende tiltak, jo mer stabilt blir driftssystemet.
Dagens ulike praksiser innen plantevern kan grupperes i tre ulike tilnærmelsesmåter (figur 1.7) som på mange måter også gjenspeiler den historiske utviklingen.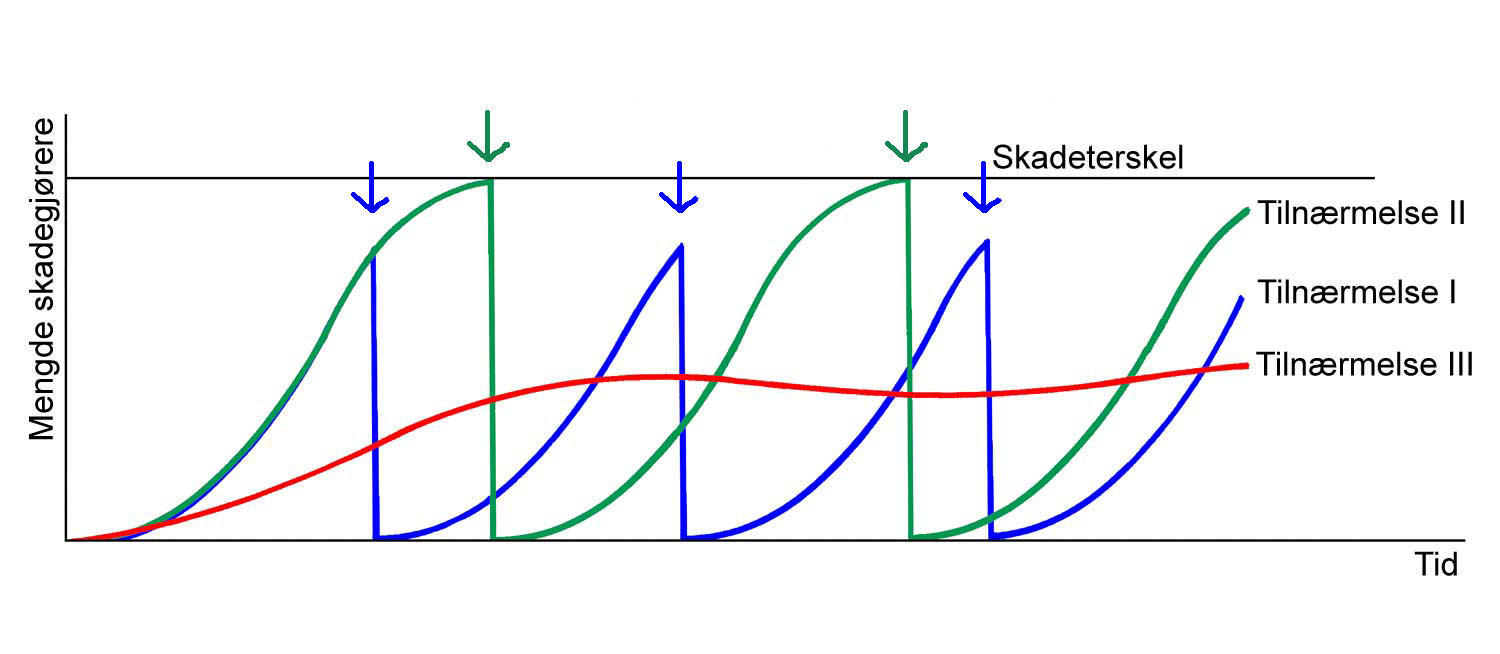
Tilnærmelse 2: Direkte tiltak gjennomføres i henhold til skadeterskel. Tilnærmelse 3: Målsetningen er at forebyggende tiltak forhindrer at skadeterskel overstiges. Pil angir når direkte tiltak gjennomføres. Tegning: Hermod Karlsen.
Tilnærmelse I. Bruk av et direkte tiltak mot en skadegjører: En har en fiende og fienden ønskes fjernet. Problemet er at denne fienden kommer tilbake fordi livsgrunnlaget for denne fortsatt er til stede. Dette betyr at det vil være nødvendig å sette inn tiltak på nytt. Denne måten å drive plantevern på kan vi si at hører til det "gammeldagse kjemikalielandbruket".
Tilnærmelse II. Aktuelle tiltak gjennomføres med henhold til bekjempelsesterskel og bruk av varslingssystemer. Dette betyr at en gjennomfører tiltak bare når det gir en økonomisk gevinst. For denne tilnærmelsen bygger konseptet fortsatt på å fjerne fienden og ikke på det å fjerne årsaken til problemet. På denne måten forsøker en å redusere uheldige bivirkninger av den opprinnelig valgte strategi.
Tilnærmelse III. Man fokuserer på forebyggende tiltak. Her blir det lagt vekt på å forhindre at problemet i det hele tatt oppstår eller bare har et begrenset og akseptabelt nivå (terskel for bekjempelse blir ikke, eller sjelden nådd). Det å løse plantevernproblemer ved hjelp av et gjennomtenkt vekstskifte hører hjemme her.
Det er stor variasjon for ulike skadegjørere om effektive forebyggende tiltak finnes. Figur 1.7 representerer, i alle fall for visse skadegjørere, mer en målsetning for framtida enn en situasjonsbeskrivelse i dag. Et eksempel kan være rognebærmøll i eple hvor effektive skadedyrmidler kan brukes (= god effekt), mens det i dag ikke finnes kjente og effektive forebyggende tiltak. På den annen side, det arbeides med å finne forebyggende tiltak også for dette plantevernproblemet. Hvis man lykkes med forebyggende plantevern, vil det kunne bety en stor gevinst fordi det øker forutsigbarheten i produksjonen, og redusere innsats og kostnader som henger samme med direkte tiltak.
Tabell 1.1 Eksempler på forebyggende og direkte plantevern 1)
| Forebyggende planteverntiltak | Direkte planteverntiltak |
| - god jord- og plantekultur | - jordarbeiding (stubbharving etc.) |
| - god grøfting | - luking |
| - optimal kalking og gjødsling | - flamming |
| - effektiv jordarbeiding | - skjerming (fiberduk) |
| - fornuftig vekstskifte | - sprøyting |
| - optimalt valg av plantearter og plantesorter (resistente sorter etc.) |
- bruk av nytteorganismer |
| - friskt frø og plantemateriale | |
| - optimal såtid, plantetid og såmengde/ planteavstand |
1) For enkelte av tiltakene som er nevnt, kan det diskuteres om de er av forebyggende eller direkte karakter
Innen generell produksjonsøkologi (tekstboks 1.7) er plantevernets rolle å kontrollere det ene hjørnet i tetraederet som vi tidligere har vist ("skadegjørere" i figur 1.1) ut fra det potensielle produksjonsnivået, avhengig av de genetiske egenskapene til kulturplantene og de abiotiske faktorene. For økologisk produksjon gjelder det spesielt å utnytte alle naturens påvirkningsmuligheter uten de "syntetiske innsatsfaktorene".
Fagområdet som omfatter studiet av hvordan ulike vekstfaktorer påvirker vekst og utvikling hos planter, er kalt produksjonsøkologi. Vi gjør oppmerksom på at begrepene som brukes i denne tekstboksen som definerende faktorer, potensielt m.fl., er oversatt fra engelsk litteratur og ikke etablerte norske utrykk. De virksomme faktorene i produksjonsøkologien kan deles i tre grupper: 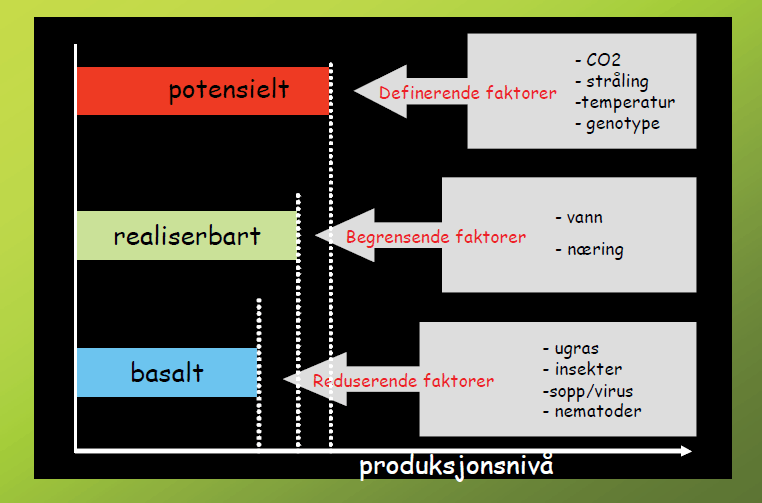
Definerende faktorer.
En plantes fotosyntese (omforming av solenergi til sukker i plantene) bestemmes av tilgang på sollys, varme og karbondioksid. Vi vet at ulike plantearter og -sorter (= genotype i figur 1.8) har forskjellig produksjonsevne, derfor er også den faktoren blant de såkalte produksjonsdefinerende faktorene. Disse faktorene bestemmer til sammen det teoretiske produksjonspotensialet («den teoretisk maksimale avling» = potensielt (rød søyle) i figur 1.8).
Begrensende faktorer.
Til denne gruppen regnes næringsstoffer og vann, og de er bestemmende for om det teoretiske produksjonspotensialet kan nås. I praksis når en svært sjelden den ideelle situasjonen som gir potensiell produksjon. I figur 1.8 er dette framstilt ved at de begrensende faktorene har redusert produksjonsnivået til «grønn søyle», som er den realiserbare delen av det teoretiske potensialet.
Reduserende faktorer.
Den tredje og siste gruppen omfatter de ulike skadegjørerne, og det blå produksjonsnivået (basalt) i figur 1.8 viser at avlingen ytterligere reduseres hvis ingen tiltak settes inn mot skadegjørerne.
Denne grupperingen av vekstfaktorene er gyldig for både konvensjonelt og økologisk landbruk.
De ulike faktorenes betydning vil variere sterkt mellom ulike driftsformer. I noen tilfeller vil for eksempel skadegjørere medføre total avlingssvikt hvis ingen mottiltak gjennomføres. Et slikt eksempel kan være at man ikke gjennomfører noe tiltak mot ugraset i en konkurransesvak kultur som gulrot.
Hvordan kan vi så bruke kunnskap fra produksjonsøkologien for å utvikle økologiske dyrkingsmetoder som gir størst mulig sikkerhet i produksjonen på både kort og lang sikt?
Litt av sammenhengene kan forklares ved å se på den definerende faktoren «genotype» i forhold til de begrensende faktorene. Ta som utgangspunkt følgende beskrivelse av etterkrigstidas landbruk:
I denne perioden har størst mulig avling vært et overordnet mål, bl.a. har dette blitt synliggjort gjennom en sterk fokusering på avling i foredlingen av nye sorter. Dette har vært mulig ved at kravet til næringstilførsel og plantevern har blitt ivaretatt gjennom tilførsel av lettløselig mineralgjødsel og bruk av syntetiske plantevernmidler. Således har man ved tilstrekkelig bruk av disse innsatsfaktorene kunnet nærme seg det potensielle avlingsnivået som er angitt i figur 1.8. Hvis man ikke tilfører disse driftsmidlene, vil produksjonen kunne komme ned på det «basale» nivået.
På kort sikt har denne framgangsmåten gitt større avlinger og redusert risikoen for avlingssvikt gjennom bruk av lettløselig mineralgjødsel og syntetiske plantevernmidler. Denne vektleggingen av avling har i mange tilfeller gått på bekostning av resistens mot skadegjørere og evne til et bedre næringsopptak ved begrenset næringstilgang. Heldigvis har resistensforedling alltid vært vektlagt, men med tilgang på effektive plantevernmidler som bakenforliggende forutsetning. Bruker man en sort (genotype) som utelukkende ivaretar målet om høyest mulig potensiell avling, men samtidig ikke har tilgang til (eller ønske om) å bruke visse typer eksterne innsatsmidler, vil produksjonen ofte vise seg å være mer risikofylt og derfor mindre avlingsstabil. Som et alternativ til beskrivelsen ovenfor, vil vi for økologisk produksjon søke følgende situasjon (figur 1.9):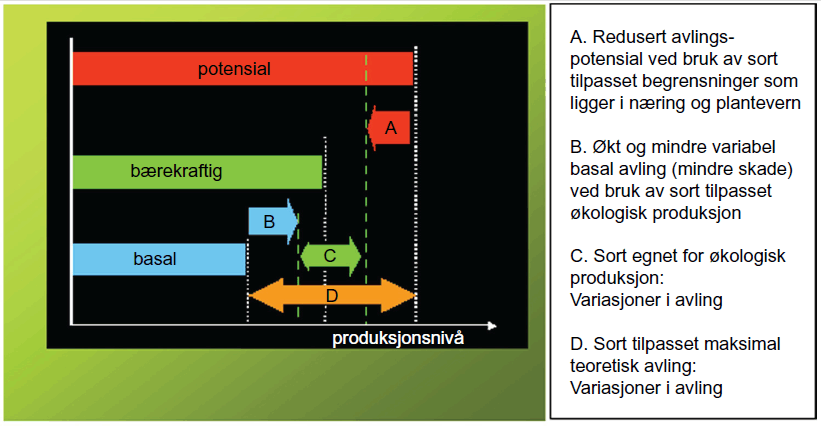
Sorter som brukes i økologisk dyrking skal gi en akseptabel avling av ønsket kvalitet, samtidig som sorten har god evne til å utnytte tilgjengelige næringsstoffer og er motstandsdyktig mot skadegjørere. Det medfører at man i praksis benytter sorter med lavere produksjonspotensial, men som samtidig er mindre avhengige av lettløselig mineralgjødsel og syntetiske plantevernmidler for å holde avlingsnivået oppe og på et stabilt nivå.
Overvåking
Regelmessig overvåking av kulturplantene er helt essensielt for å oppdage angrep tidlig. Dette er like viktig i en åpenåkerkultur som i et veksthus. Kontroller bør foregå hyppig, og tettest når man av erfaring eller ved varsling vet at angrep kan forekomme. Overvåking og registrering for å danne seg et riktig skadegjørerbilde er like viktig for både ugras, sopp og skadedyr. Den visuelle kontrollen, med bruk av feller for å se om for eksempel gulrotflua forekommer, er avgjørende for denne typen skadegjørere. For kontroll av visse insekter er det også viktig at man ikke bare ser i kantene, men går ut i åkeren for å se over hele feltet. Det samme gjelder ugras. Vindspredning av insekter kan gi forekomst på spesielle steder og vegetativ formering av rotugras fører til "ugrasroser".
Skadeterskler
Begrepet skadeterskel blir mye brukt i sammenheng med direkte tiltak mot skadegjørere.
Begrepet er utviklet og gitt innhold i et mer omfattende kunnskapsområde som fagfolk ofte benevner skadeterskelteori. Som omtalt før, står skade, avlingstap og økonomisk tap i sammenheng med hverandre, men sammenhengen er ikke statisk (fastlåst). Det finnes to forskjellige skadeterskelbegrep. Bekjempelsesterskelen defineres som det laveste nivået av en skadegjører som krever innsats for å unngå avlingstap. Den økonomiske skadeterskelen forteller oss når det er lønnsomt å sette inn tiltak mot skadegjøreren, dvs. skadeomfanget veid opp mot kostnadene man har ved gjennomføring av tiltak.
Mange skadegjørere er vanskelige å vurdere mht. hvordan angrepet vil utvikle seg, bl.a. fordi mange skadegjørereres utvikling er relatert til vær og klimaforhold. Verken bekjempingsterskel eller økonomisk skadeterskel er altså fastlåste verdier. De er avhengig av framtidig utvikling av en skadegjører, og kan dermed være mer eller mindre usikre.
Bøndenes egen erfaring, kombinert med en forståelse av hvordan de enkelte skadegjørere kan utvikle seg under visse forhold, hjelper sterkt til å bedømme de aktuelle og framtidige plantevernproblemene på gården. Dette gjelder også tolkingen av skadeterskelnivået.
Prognoser og varsling
Prognoser for angrep og varsling av mulig skade er etablert for en del skadegjørere i viktige jord- og hagebruksvekster (tekstboks 1.8).
Varsling Innen PlanteSkadegjørere (VIPS) er en gratis varslingstjeneste på Internett om angrep og skader av sjukdommer, skadedyr og ugras i viktige jord- og hagebruksvekster. Tjenesten er et tiltak under Handlingsplan for redusert risiko ved bruk av plantevernmiddel og er utviklet av Bioforsk og Landbrukets Forsøksringer (LFR).
VIPS er fritt tilgjengelig for alle på www.vips-landbruk.no og gir informasjon til dyrkere og veiledere om fare for angrep eller ikke og dermed behov/ikke behov for sprøyting, eventuelt andre tiltak. Varsling er en viktig del av integrert plantevern og er et hjelpemiddel for å unngå unødvendig bruk av plantevernmidler og dermed unngå ekstra kostnader for dyrkere og unødvendig miljøbelastning. I tillegg til varsling har nettstedet også overvåking av skadegjørere ved at det legges ut meldinger om førstefunn og utvikling/skadeomfang av en rekke sjukdommer og skadedyr i ulike distrikter.
VIPS omfatter skadegjørere i korn, potet, grønnsaker og frukt (se tabell under). Varslene blir laget på grunnlag av klimadata, værprognoser, observasjoner av angrep i felt, skadeterskler og modeller for utvikling av skadegjørere/vertplanter.
Klimadata levers av ca. 75 værstasjoner (Bioforsk eier de fleste, noen er private/eies av forsøksringer) som er plassert i de viktigste dyrkingsområdene. Værprognosene leveres av Meteorologisk Institutt. Forsøksringene registrerer angrep av skadegjørere i felt og data legges inn i modeller (utvikla av forskere ved Bioforsk) for den enkelte skadegjører. Varslene vises på internett ved hjelp av fargesymboler: Rød betyr infeksjonsfare, gul betyr mulig infeksjonsfare og grønn betyr ingen infeksjonsfare.
Varsler i VIPS (per 2005)
| Korn | Potet og grønnsaker | Frukt og bær |
| Grå øyeflekk | Potettørråte | Epleskurv |
| Hveteaksprikk | Kålflue | Eplevikler |
| Byggbrunflekk | Kålfly | Rognebærmøll |
| Gulrotflue | Gråskimmel i jordbær (testversjon) |
Brukere kan registrere og legge inn egne observasjoner/forutsetninger/tiltakpå egen gård og motta personlig varsel. Ved vurdering av behov for å sette i verk eventuelle tiltak kan det være lurt å kontakte den lokale veiledningstjenesten. Ringlederen sitter inne med stor kunnskap om lokale vekstforhold, og kan derfor «tolke» det lokale varslet med tanke på egen gård og produksjon.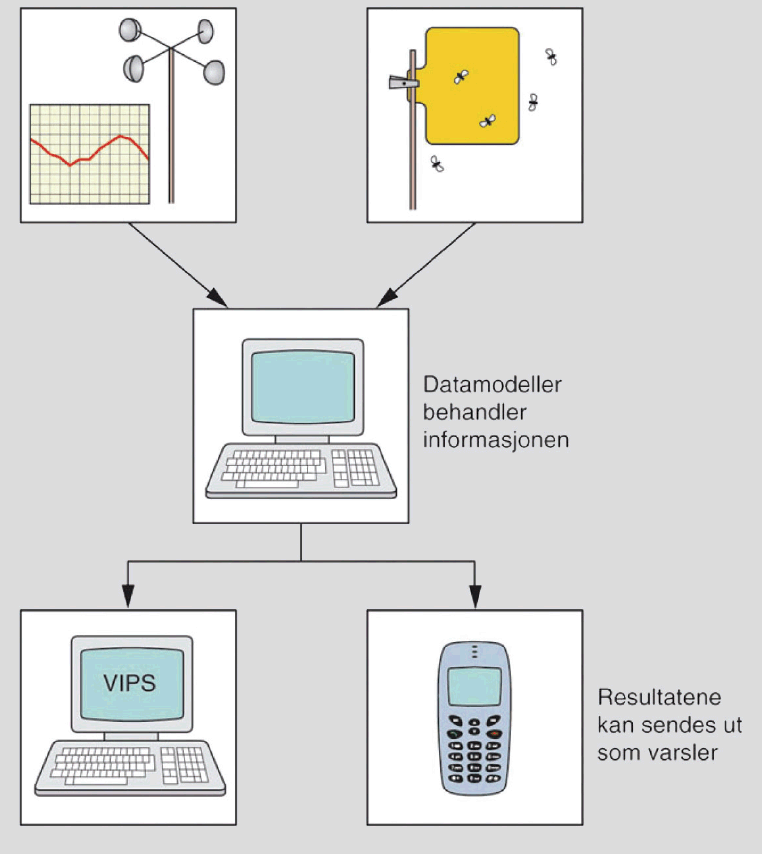
VIPS ugras i korn er et nettbasert hjelpemiddel for å beregne behandlingsbehov ved ugrasangrep i kornåker. Dette er en norsk versjon av det danske Plantevern Online. Programmet gir ikke varsel om ugras, ugraset kommer uansett, men det gir råd om middelvalg og dose på bakgrunn av opplysninger om kornart, forventet avlingsnivå, temperatur, jordtype og ugrasarter/mengder/utviklingsstadium, med eller uten gjenlegg.
Mange vil sikkert, og det med god grunn, stille spørsmålet om relevansen av VIPS i forhold til økologisk landbruk. Bruk av varsler og skadeterskler er bygd på at man har tilgang på direkte tiltak når problemer oppstår, og det har man jo ofte ikke ved økologisk dyrking. Dette kan endre seg i fremtiden for eksempel ved at planteekstrakter etc., godkjennes mot visse skadegjørere. Ellers vet vi for eksempel at det finnes gulrotdyrkere i dag som legger på fiberduk ved varsling av gulrotflue.
IFOAM 2004: The World of Organic Agriculture. Statistics and Emerging Trends 2004 (Red. Miller, H. og M. Yussefi). 167 s.
Källander, I. 1989: Växföljden - grunden i odlingssystemet. I: Jordbruksbok för alternativ odlare. Stockholm, s. 184-194
Landbruksdepartementet 1999: Stortingsmelding nr. 19, 1999-2000: Om norsk landbruk og matproduksjon.161 s.
Porceddu, E. og R. Rabbinge, 1997: Role of research and education in the development of agriculture in Europe. I: M.K. van Ittersum og S.C. Geijn (Red.) Perspectives for Agronomy - Adopting Ecological Principles and Managing Resource Use. Pages 3-15 (in series Developments in Crop Science 25. Elsevier Science.
Ruissen, T. 2002: Plantevern i økologisk landbruk. I: Økologisk landbruk. NORSØK-rapport 1-2002, s. 31-38
Serikstad, G.L. 2000: Økologisk landbruk. Småskrift (NORSØK) nr. 2 / 2000
Skjelvåg, A.O. 1992: Bærekraftige produksjonssystem. Forelesningsnotat i hovedkurs i plantekultur ved Norges Landbrukshøgskole.
www.debio.no
www.mattilsynet.no
www.bioforsk.no
www.hihm.no
www.vips-landbruk.no
Kunnskap om skadegjørernes biologi er viktig både for å unngå at plantevernproblemer oppstår, og for å kunne gjennomføre direkte mottiltak i kulturene på en best mulig måte. Derfor vil vi i dette kapitlet komme relativt grundig inn på ulike forhold omkring ugrasets, skadedyrenes og sjukdomsorganismenes evne til å overleve sammen med plantene vi dyrker. Med et fint ord kan vi kalle denne evna for deres «livsstrategier». Vi vil i dette kapitlet komme mer detaljert inn på ugrasets enn skadedyrenes og sjukdomsorganismenes livsstrategier. Dette blir gjort fordi ugraset generelt sett er mer universelt ved at de ulike artene finnes i de fleste av våre kulturvekster, mens mange skadedyr og sjukdomsorganismer er mer spesifikke i valg av kultur eller vertsplante. Livssykluser og livsstrategier for mange skadedyr og sjukdomsorganismener, vil derfor bli nærmere omtalt i Bind 2, 3 og 4 i forbindelse med de ulike kulturvekstene.
Det er delte meninger om hvilke ord en skal bruke på kampen mot skadegjørerne. For eksempel vil mange oppfatte betegnelsen ugrasbekjempelse mer eller mindre synonymt med totalbekjempelse, oftest i forbindelse med bruk av ugrasmidler (herbicider). Noen mener derfor at dette ordet ikke hører hjemme i et mer økologisk perspektiv. Begreper som ugrasregulering, ugraskontroll og ugrastiltak er mer nøytrale ord. For alle skadegjørergrupper vil de ulike tiltakene kunne deles inn slik:
Forebyggende tiltak vil for alle grupper av skadegjørere være en bærebjelke i økologisk plantevern. Ofte vil det i tillegg være nødvendig med direkte tiltak for å forhindre videre utvikling når en skadegjører en gang har slått til i en kultur. Å vite noe om både skadegjørernes livssyklus og biologi og deres samspill med miljøet omkring, det vi i sum kan kalle deres økofysiologi, er avgjørende for at de forebyggende tiltakene vi setter inn skal fungere best mulig.
I de fleste sammenhenger blir ugras betraktet som viltvoksende planter som vokser blant kulturplantene, og som konkurrerer med disse om vekstfaktorer som lys, vann, næring og plass. Begrepet ugras defineres gjerne på forskjellige måter, men her nevner vi bare en kort, generell definisjon: Ugras er uønska planter, d.v.s. planter som opptrer på en plass der de ikke er ønsket med hensyn til plassens bruksområde (se også Korsmos definisjon i tekstboks 2.1).

Emil Korsmo ble født i 1863, og døde i 1953, vel 90 år gammel. Han var en pionér innen ugraslære (herbologi), og gjorde kampen mot ugraset til sin livsoppgave. Han fungerte som statskonsulent i ugrasspørsmål i årene 1913-20, og som professor i herbologi ved Norges Landbrukshøgskole (nå: Universitetet for miljø- og biovitenskap) i årene 1920-33. Korsmo utgav en rekke bøker og skrifter, for eksempel Ugress i nutidens jordbruk (1925, ny utgave i 1954), Ugressplancher (1918, ny utgave 1934-38), plansjeverket Ugressfrø med tekst og register på 11 språk (1935) og et stort, illustrert verk om ugrasplantenes anatomi, Anatomy of Weeds (1954). Ugrasplansjene er senere blitt utgitt i bokform (1981, 1986 og 2001) av Landbruksforlaget. 5 nye arter ble tatt inn i boken i 2001-utgaven, der grafikken er utført av tegneren Hermod Karlsen. Flere av verkene til Korsmo er oversatt til andre språk. Allerede i sin første bok, Ugræs i Ager og Eng som utkom i 1896, definerte han ugras på følgende måte: «Ved ugræs forstår man alle de paa dyrket mark opptrædende planter som man ikke tilsigter at have der ... » Korsmo var den første forsker som inndelte uønskete plantearter i biologiske grupper. Formålet var den gang som nå: sikker bestemmelse og bekjempelse av ugrasene. Ved ulike forsøk som ble utført, kunne effektiv ugrasbekjempelse gi 20-40 % avlingsøkning. Noen av bekjempingsmetodene Korsmo beskrev, som luking og harving, er fremdeles aktuelle, mens de kjemiske metodene ved bruk av svovelsyre, salpetersyre og kalsiumcyanid, hører historien til. Ett unntak er jernsulfat, som fremdeles er i bruk til kjemisk bekjempelse av mose i plen.
Dette understreker at svaret på spørsmålet om en plante skal karakteriseres som ugras, d.v.s. uønsket, eller ikke, avhenger av situasjonen. Ingen planteart er alltid ugras, men mange arter kan i gitte situasjoner opptre som ugras. Noen arter gjør dette svært ofte, andre langt sjeldnere. Det er derfor feil å si at ugras gjør nytte. Ugras gjør skade. Når de samme artene i andre situasjoner er nyttige, er de ikke uønsket, og følgelig heller ikke ugras. Figur 2.1 oppsummerer en del av den "dobbeltrollen" som plantene vi her diskuterer, kan ha når de opptrer som henholdsvis ugras og nytteplanter.
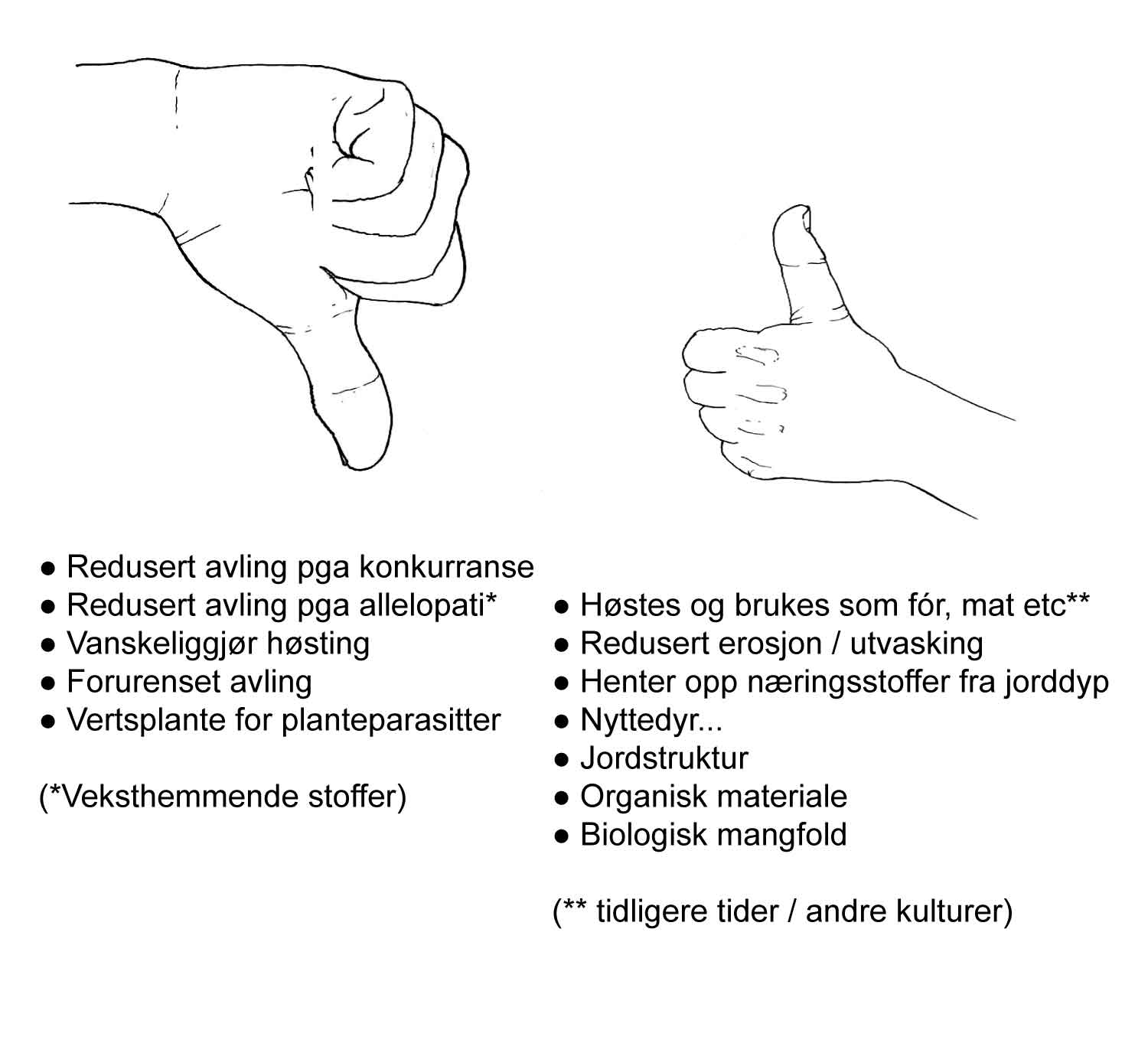
I noen tilfeller kan til og med kulturplantene selv opptre som ugras. Dersom en oljevekst er dyrket ett år, kan spillfrø overvintre og føre til oppblomstring i neste kultur som for eksempel kan være korn. Et annet eksempel er dyrking av ulike kornarter etter hverandre i et omløp. Spillkorn fra havre det ene året kan gjøre seg gjeldende som ugras i bygg året etter.
For å forstå ugrasfloraens sammensetning i dagens jord- og hagebruk, er det nyttig å kjenne noe til de enkelte planteartenes innvandringshistorie. Under den siste istida var hele Skandinavia, med unntak av noen fjelltopper (nunataker) og trolig en smal kyststripe, dekket av is. En regner med at Skandinavia utelukkende har vært mottakerland for planter etter istida, bortsett fra noen overlevende fjellplanter. De fleste artene, ugrasene inkludert, har vandret inn fra sør og sørøst, via Mellom-Europa og Danmark. Noen av dagens ugras (bl.a. vassarve, burot og ulike meldearter) var blant de tidligste pionerplanter i kjølvannet etter avsmeltingen.
En kan skille mellom de artene som vandret inn til landet ved egen hjelp, de såkalte spontane eller heimlige artene, og de artene som ble brakt til landet ved hjelp av mennesket, tilsiktet eller utilsiktet. Den siste gruppen arter kalles for antropokore eller innførte arter, og utgjør nå nesten halvparten av alle arter. Av totalt ca. 2400 plantearter i Norge, er det ca. 250 arter, eller noe over 10 %, som gjør seg gjeldende som ugras. Ca. 100 av disse artene er vanlige over mer eller mindre hele landet.
På grunnlag av fossile funn av frø og andre plantedeler av ulik størrelse (makrofossiler), funnet i naturlige avleiringer i vann (for eksempel bunnslam i innsjøer, leire og torv), har en kunnet datere ved hjelp av C-14-metoden (radioaktivt karbon) den tidligste innvandringen av planter i perioden ca. 13 000-3000 f.kr. Da åkerbruket begynte ca. 3000 f.kr. i den yngre steinalder og fram til og med vikingtida (ca. 1050 e.kr.), er de vesentlige funn fra avtrykk i kar, potteskår og leire. Senere skriver funnene seg fra for eksempel jord, stolpehull, hus, lagre, latriner og humusholdige lag. Innvandringshistorien er også basert på analyse av pollen eller blomsterstøv, gjerne kalt mikrofossiler.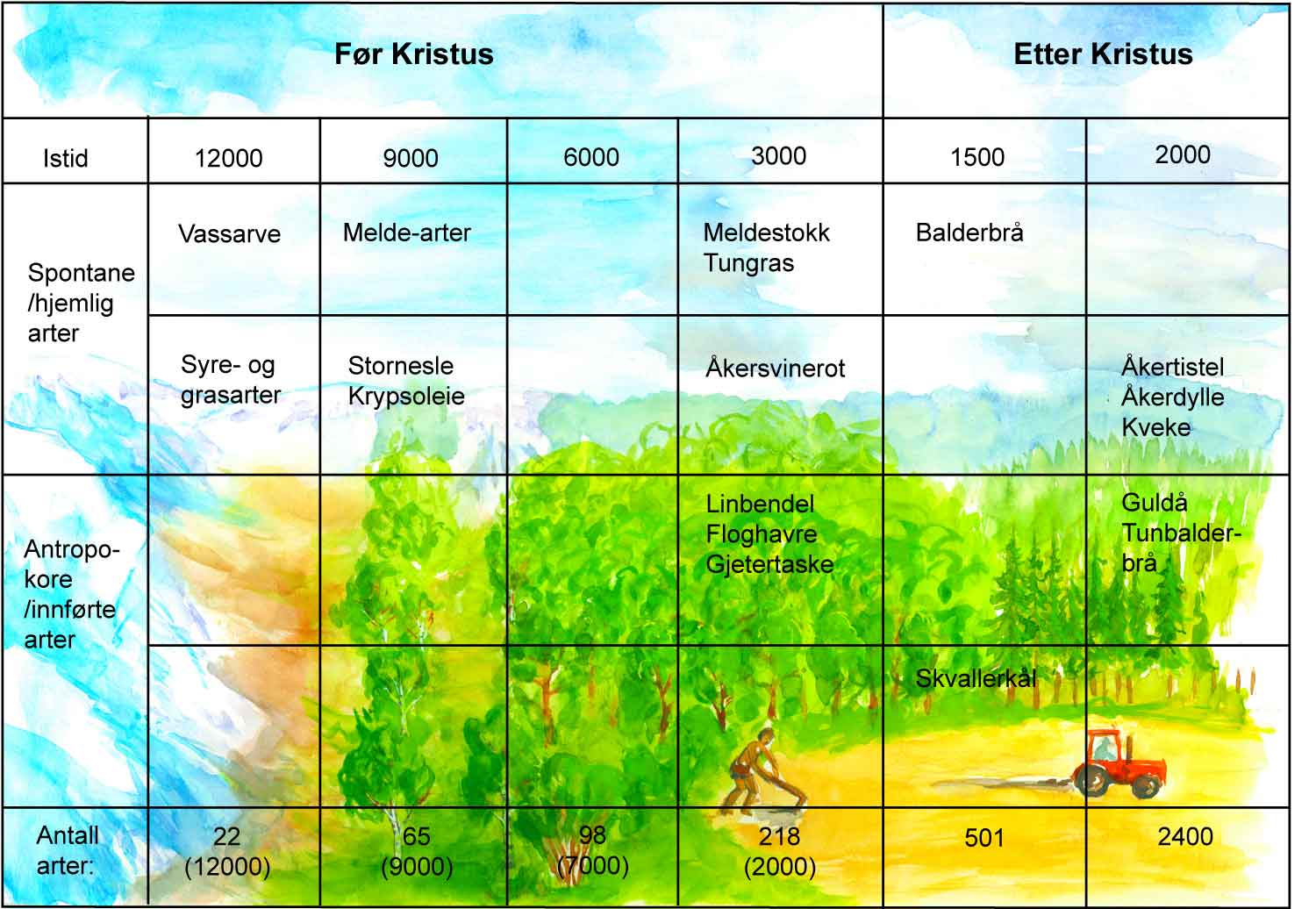
Figur 2.2 viser når en har kunnet datere de første eksemplar av arter som i dag ofte opptrer som ugras i Skandinavia. Både klimaet og vegetasjonen har gjennomgått store endringer i de over 15 000 årene som har gått siden istida, samtidig som landet hevet seg. Botanikeren Axel Blytt mente å kunne påvise (i 1876) to store skogperioder på grunnlag av to ulike stubbelag i myrer på Vestlandet. Da må en regne med at de fleste urteplanter var trengt langt tilbake. Det nye åkerbruket og fedriften førte med seg betydelige forandringer i floraen. Da det ble ryddet i de tette urskogene, ble landskapet etter hvert mer og mer åpent. Særlig for lyselskende arter hadde edelløvskogene vært en innvandringsbarriere. Nå fant disse artene store muligheter i de mange skogkantene som oppsto. Også i våre dager er skogkantene viktige lokaliteter for lyselskende busker og urter. Jordbrukskulturen førte til en renessanse for en rekke arter som hadde spilt en betydelig rolle i den første koloniseringen av landområder, men som hadde fått redusert sine muligheter etter hvert som skogene overtok. Gjødsling av åkrer begunstiget bl.a. nitrofile (nitrogenelskende) strandplanter som svinemelde, meldestokk, klengemaure, åkertistel og åkerdylle til å slå seg ned der. Dessuten brakte mennesket etter hvert med seg mange nye arter sammen med såkorn og grasfrø.
Ulike dyrkningsmessige forhold har, fra det tidlige svedjejordbruket da vegetasjonen ble brent ned før dyrking, og som varte til ut på 1800-tallet og fram til dagens industrijordbruk, endret seg betydelig og medførte at ugrasfloraen gjennomgikk store endringer både med hensyn til hvilke arter og antallet arter som fantes. Et eksempel kan være floghavrens fluktuasjoner gjennom 17-, 18- og 1900-tallet (figur 2.3).
På 1700-tallet var det mange steder en relativt ensidig korndyrking, som medførte store problemer med floghavre. Dette systemet ble på 1800-tallet etterfulgt av et mer mangfoldig vekstskifte med flerårig eng, noe som langt på vei eliminerte problemene med floghavre. I dagens moderne jordbruk, hvor korn ofte dyrkes ensidig, ser vi at problemet med denne arten har kommet tilbake. Dagens moderne landbruk har også medført at antall problematiske ugras har gått tilbake. I eldre tider med "svakere" kulturvekster og mindre effektiv jordarbeiding, kunne mange arter etablere seg, mens dyrkingsmiljøet i dag passer for færre arter. Undersøkelser fra Sverige viser at i dag utgjør de 10 vanligste artene om lag 75 % av ugrasbiomassen, mens for 30 år siden måtte det 20 arter til for tilsvarende biomasse.
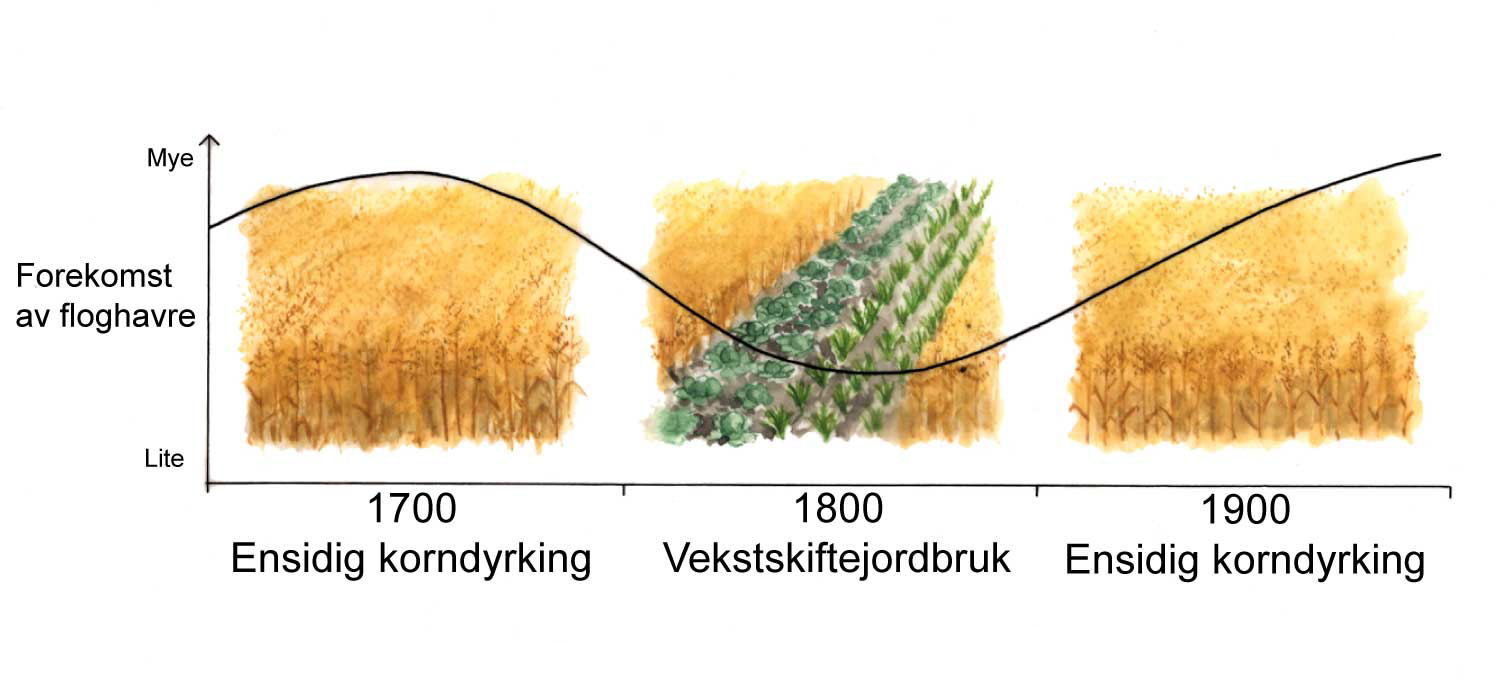
Mulighetene for en art til å konkurrere med kulturveksten og formere seg, er avhengig av ugrasplantens vokseegenskaper i forhold til kulturplanten. Særlig er den årlige vekstrytmen viktig, eksempelvis spiretidspunktet, og om og når jordarbeiding blir utført i løpet av året. Både ugraset og kulturen, inkludert alle dyrkingstekniske tiltak bonden utfører, inngår i agroøkosystemet. I ugraslæren (herbologien) deler man gjerne ugraset i biologiske grupper etter de egenskapene som har størst praktisk interesse, uten omsyn til den vanlige botaniske systematikken. Av særlig interesse i denne sammenhengen er levealderen og formeringsmåten til ugraset. Denne inndelingen ble utførlig beskrevet av Korsmo (tekstboks 2.1)
Emil Korsmo ble født i 1863, og døde i 1953, vel 90 år gammel. Han var en pionér innen ugraslære (herbologi), og gjorde kampen mot ugraset til sin livsoppgave. Han fungerte som statskonsulent i ugrasspørsmål i årene 1913-20, og som professor i herbologi ved Norges Landbrukshøgskole (nå: Universitetet for miljø- og biovitenskap) i årene 1920-33. Korsmo utgav en rekke bøker og skrifter, for eksempel Ugress i nutidens jordbruk (1925, ny utgave i 1954), Ugressplancher (1918, ny utgave 1934-38), plansjeverket Ugressfrø med tekst og register på 11 språk (1935) og et stort, illustrert verk om ugrasplantenes anatomi, Anatomy of Weeds (1954). Ugrasplansjene er senere blitt utgitt i bokform (1981, 1986 og 2001) av Landbruksforlaget. 5 nye arter ble tatt inn i boken i 2001-utgaven, der grafikken er utført av tegneren Hermod Karlsen. Flere av verkene til Korsmo er oversatt til andre språk. Allerede i sin første bok, Ugræs i Ager og Eng som utkom i 1896, definerte han ugras på følgende måte: «Ved ugræs forstår man alle de paa dyrket mark opptrædende planter som man ikke tilsigter at have der ... » Korsmo var den første forsker som inndelte uønskete plantearter i biologiske grupper. Formålet var den gang som nå: sikker bestemmelse og bekjempelse av ugrasene. Ved ulike forsøk som ble utført, kunne effektiv ugrasbekjempelse gi 20-40 % avlingsøkning. Noen av bekjempingsmetodene Korsmo beskrev, som luking og harving, er fremdeles aktuelle, mens de kjemiske metodene ved bruk av svovelsyre, salpetersyre og kalsiumcyanid, hører historien til. Ett unntak er jernsulfat, som fremdeles er i bruk til kjemisk bekjempelse av mose i plen.
Sommerettårige ugras
Sommerettårige arter lever bare en sommer. De spirer opp av frø om våren, blomstrer og setter frø. Deretter dør hele planten, inklusiv roten (figur 2.4). Disse artene overvintrer altså bare som frø. Frøproduksjonen er som regel svært rikelig, og frøet modner samtidig med eller før den kulturveksten ugraset vokser i. Det frøet som faller på jorda, spirer vanligvis først neste vår. Skulle frøet spire alt samme høst, vil planten som regel fryse i hjel i løpet av vinteren. I særlig milde vintre kan visse arter greie seg. Blir frøet gravd dypt ned under jordarbeidinga, kan det ligge i jorda i mange år uten å miste spireevna.
Sommerettårige ugras kan bare utvikle seg i større mengder der jorda blir arbeidet om våren. De fleste av våre vanligste ugras i åker og hage hører til denne gruppen. De viktigste artene er: Floghavre, meldestokk (tekstboks 2.2 og figur 2.4), kvassdå, guldå, linbendel, hønsegrasartene, tungras, vindeslirekne, åkergull, åkersennep, åkerkål, åkerreddik, klengemaure, åkerstemorsblom, tunbalderbrå, åkervortemjølk, hønsehirse og jordrøyk.
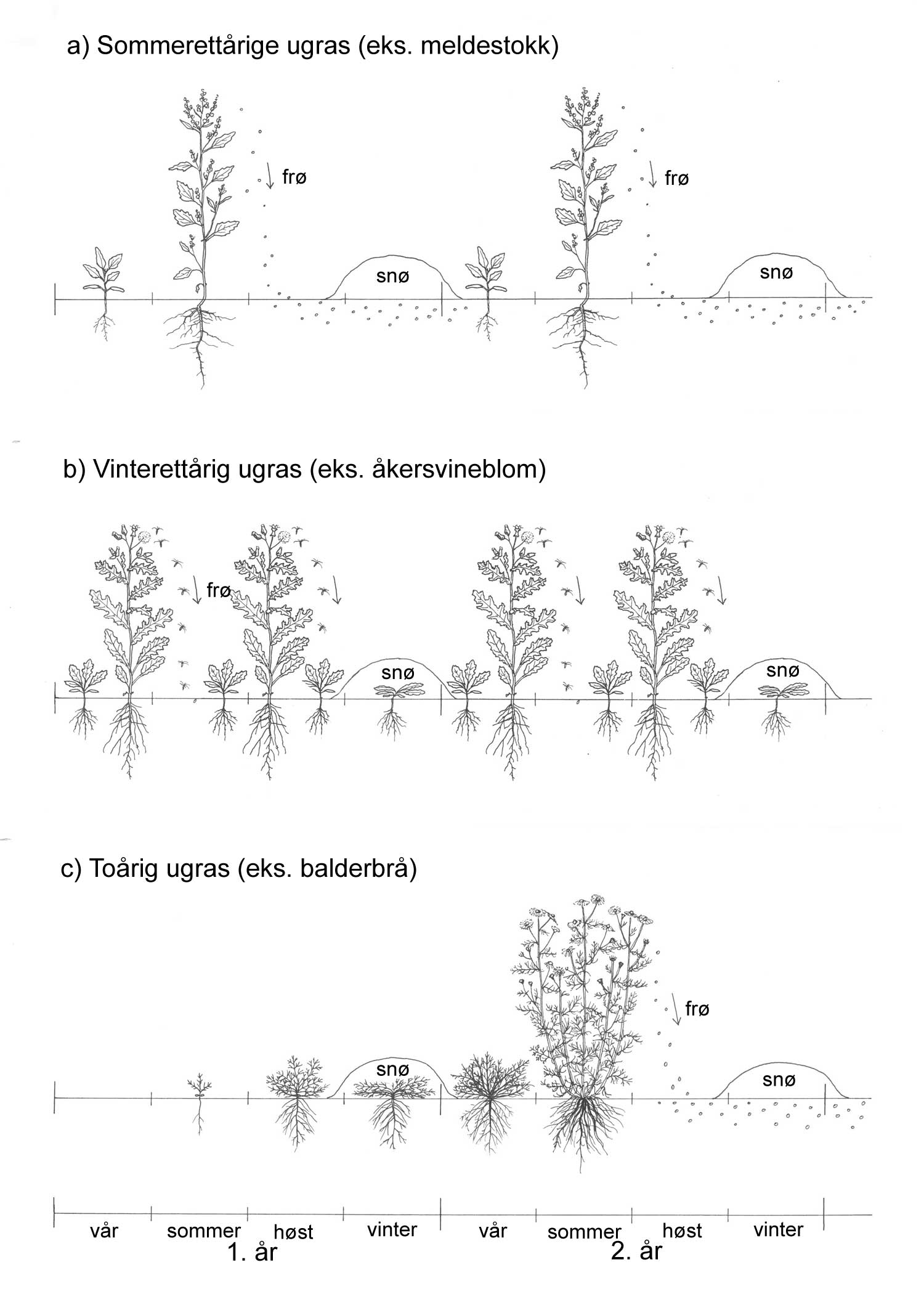
Vinterettårige ugras
Vinterettårige arter har normalt evne til å overvintre. Frøet kan spire gjennom hele sommerhalvåret. Spirer det tidlig nok, blomstrer plantene og setter modent frø på samme måten som de sommerettårige. Disse frøene kan igjen spire og utvikle nye frøproduserende planter. Spirer frøet så sent i vokseperioden at planten ikke når full utvikling, overvintrer den, blomstrer og setter frø neste vår eller sommer (figur 2.4). Frøene fra disse plantene kan så spire, og siden det ennå er tidlig på året, har plantene gode muligheter til å produsere egne frø før vinteren. Dermed kan en oppnå to frøgenerasjoner på ett år. I noe varmere land enn Norge, for eksempel England, kan en til og med få tre frøgenerasjoner i året.
Vinterettårige ugras er, som vi skjønner, mer allsidige enn de sommerettårige. De vokser derfor godt både i vårsådde og i høstsådde kulturer, men som de sommerettårige artene, er også de vinterettårige avhengige av løs jord for å kunne utvikle seg i større omfang. Vi har bare ni vinterettårige ugras som er særlig viktige: Vassarve, gjetertaske, rødtvetann, pengeurt, åkersvineblom (tekstboks 2.3 og figur 2.4), haremat, tunrapp, stemorsblom og åkerminneblom.
Denne informasjonen finnes i artikkelen om Åkersvineblom i leksikonet
Denne informasjonen finnes i artikkelen om Balderbrå i leksikonet
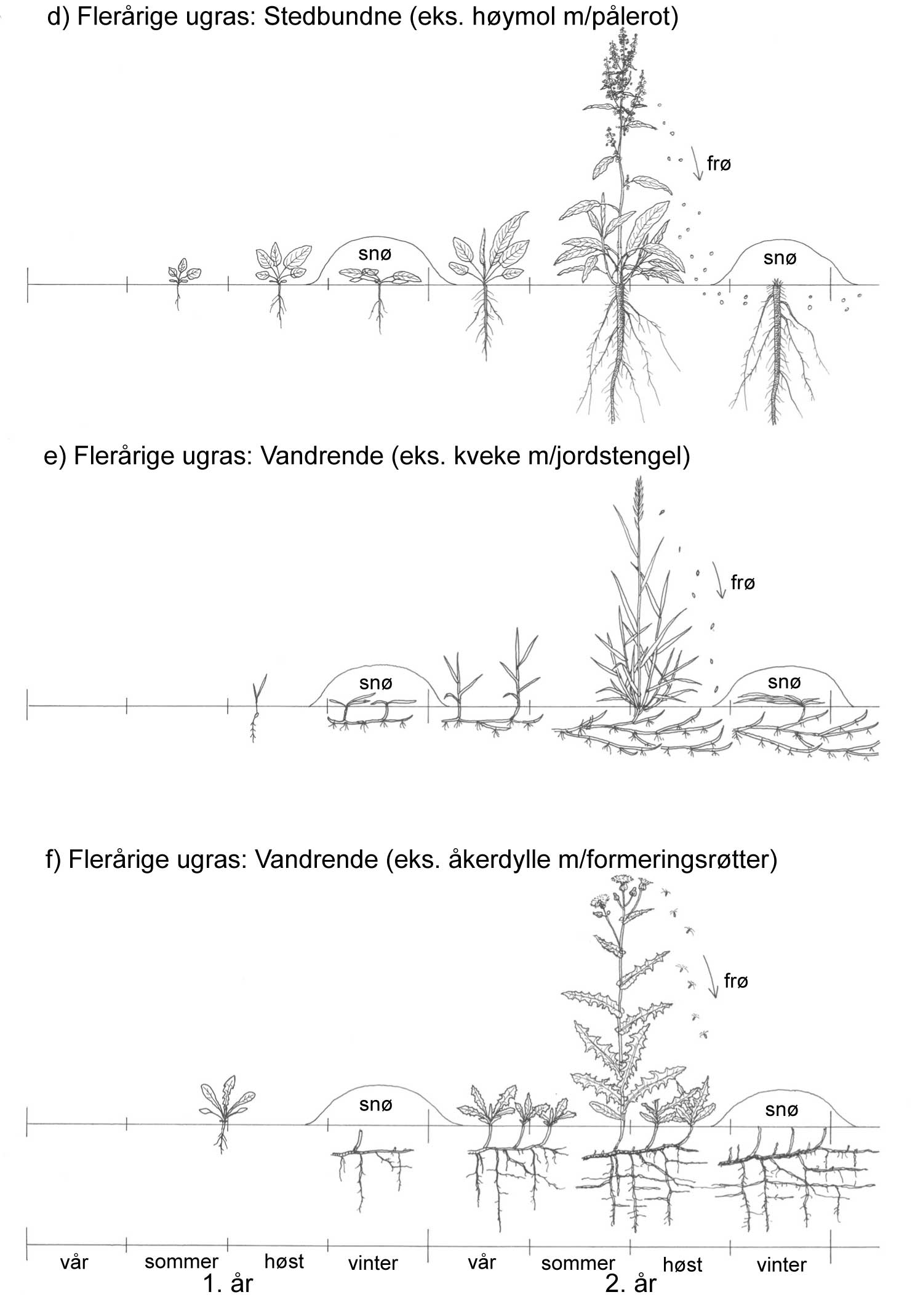
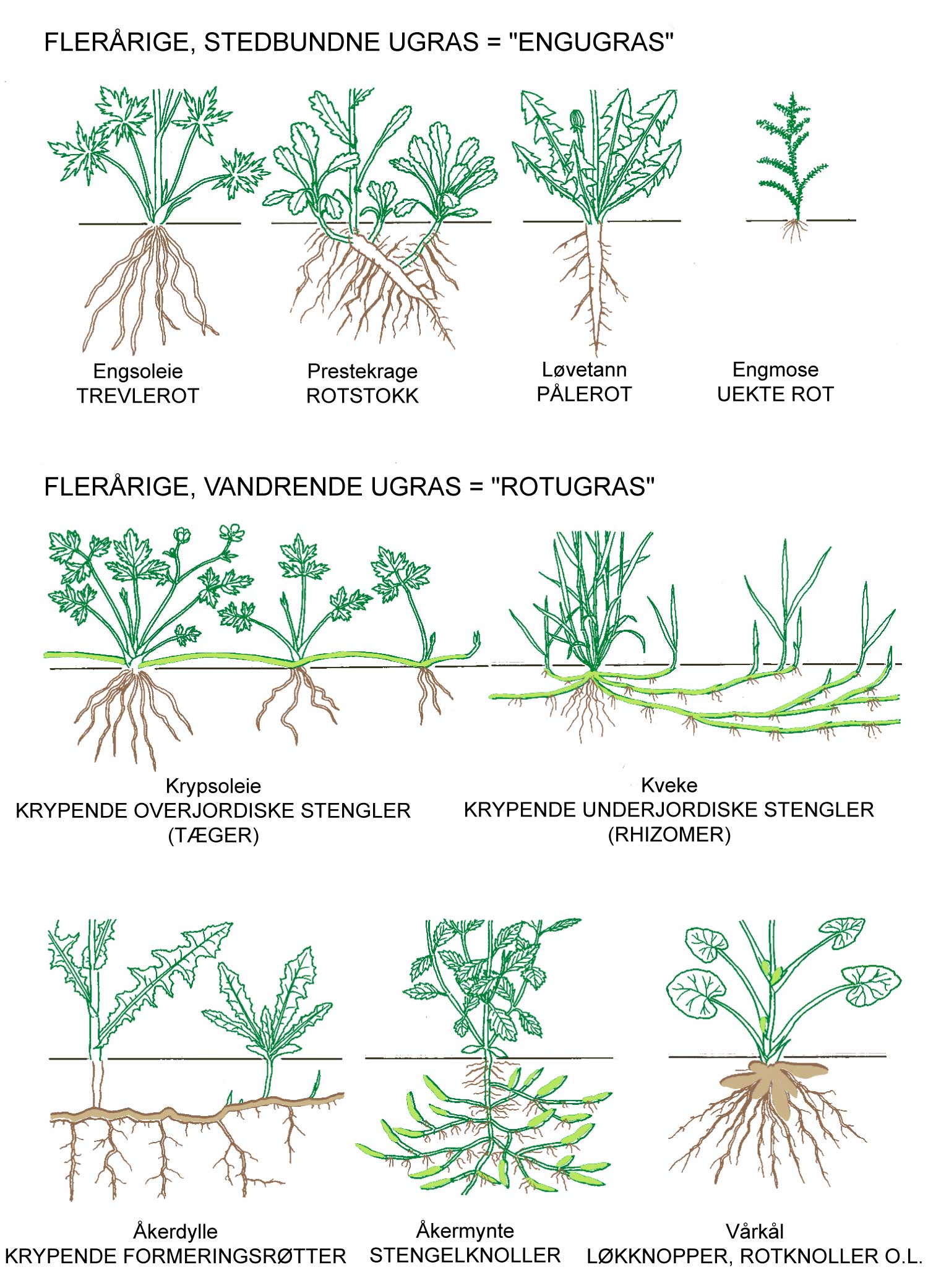
Det er særlig i eng og beite, og andre steder der planten kan vokse i fred i lengre tid, at vi finner ugras som hører til gruppen stedbundne ugras. De blir derfor ofte kalt «engugras».
Denne ugrasgruppen kan ellers deles i fire undergrupper etter rottype: trevlerot, rotstokk, pålerot og uekte rot (figur 2.9). De viktigste artene i de fire undergruppene er:
Denne informasjonen finnes i artikkelen om Vanlig høymole i leksikonet
Uekte rot betyr at «roten» ikke er bygd for næringsopptak som hos høyere planter/karplanter, men bare tjener som festeorgan, som hos tang og tare i sjøen. Næringsopptak i moser (og i tang og tare) skjer direkte via bladoverflata.
Vandrende ugras
Alle arter i gruppen vandrende ugras har kontinuerlig, vegetativ formering og spredning. De formerer seg dessuten med frø eller sporer. Når de vokser opp fra frø, lager de i spiringsåret bare bladrosett og rot som overvintrer. De fleste artene blomstrer og setter frø første gangen året etter, altså i 2. leveåret (figur 2.8), men noen først 3. året (hestehov, hundekjeks og skvallerkål). Mange arter er svært frørike. Vandrende ugras har altså evne til å spre seg vegetativt, uten ytre inngrep. I dagligtale blir de ofte kalt «rotugras». Noen inkluderer gjerne også de stedbundne i dette begrepet. Mange av de mest brysomme ugrasene, både i åker og grasmark, hører til denne gruppen.
Etter den vegetative formeringsmåten kan rotugraset deles i fem undergrupper: Krypende, rotslående stengler, krypende jordstengler, krypende formeringsrøtter, stengelknoller i jorda, annen vegetativ formering (figur 2.9). De viktigste artene i de fem undergruppene er:
Denne informasjonen finnes i artikkelen om Kveke i leksikonet
Denne informasjonen finnes i artikkelen om Åkerdylle i leksikonet
Denne informasjonen finnes i artikkelen om Åkertistel i leksikonet
Ugrasklokke har for eksempel både krypende jordstengler og pålerøtter som vokser ut fra jordstenglene. Hundekjeks har en form for oppsplitting av øvre del av røttene, og er således «svakt» vandrende.
Forekomst av ugras i ulike kulturer
Som figur 3.28 viser, er det stor forskjell i konkurranseevne mellom de ulike kulturvekster. Dette fører til at hvis vi ikke setter inn direkte tiltak mot ugraset, kan vi forvente at det blir langt mer ugras i for eksempel gulrot eller kål enn i kornvekstene. Et annet spørsmål er om vi vil finne de samme ugrasartene i alle kulturvekster, og om de samme artene dukker opp uavhengig av så- og plantetidspunkt. Finner vi eksempelvis de samme artene i vårkorn og høstkorn?
Korsmo, som altså først etablerte en biologisk inndeling av ugraset, utnyttet ikke muligheten for systematisering av hvilke arter som etablerer seg i de ulike kulturvekster. Likevel har hans detaljerte beskrivelser utvilsomt gitt viktig informasjon som underlag for bedømming av hvilke ugras som opptrer i ulike kulturvekster. Den svenske ugrasforskeren Sigurd Håkansson har kombinert Korsmos biologiske inndeling med grupper av ulike kulturvekster, basert på aktuell litteratur og intervjuer av biologer og dyrkere, og bygd opp en matrise som viser hvilke plantearter/plantegrupper som mest sannsynlig vil kunne opptre som ugras og skape problemer i ulike kulturvekster (tabell 2.1). De sommerettårige artene etablerer seg først og fremst i kulturvekster som også er sommerettårige, eksempelvis poteter, grønnsaker, vårkorn og våroljevekster. I disse kulturene vil naturligvis vårspirte, vinterettårige arter også etablere seg (jfr. åkersvineblom, figur 2.6). Ved dyrking av høstsådd korn og oljevekster, vil det være mindre av sommerettårige arter, men desto mer av vinterettårige og toårige ugras. Av flerårige arter, vil stedbundne og vandrende arter som er følsomme for jordarbeiding, hovedsakelig finnes i enga. De mest problematiske flerårige artene, helst arter med underjordiske stengelutløpere som er motstandsdyktige mot jordarbeiding, og arter med rotutløpere, kan forventes å finnes i nær sagt alle kulturvekster. Flerårig eng kan virke sanerende på flerårige arter med rotutløpere, for eksempel åkertistel.
Tabell 2.1 Forekomst av ulike biologiske ugrasgrupper i de forskjellige kulturvekster (modifisert etter Håkansson 1995)
| Biologiske ugrasgrupper | Ettårige kulturvekster | Eng | |||||
| Grønnsaker og poteter |
Vårkorn og oljevekster |
Høstkorn og oljevekster |
Yngre | Eldre | |||
| Ettårige arter: | |||||||
| Sommerettårige | +++ | +++ | +/++ | + | 0 | ||
| Vinterettårige | +++ | ++ | +++ | ++ | + | ||
| Toårige arter* | 0 | 0 | ++ | + | 0 | ||
| Flerårige stedbundne arter | + | 0 | 0 | ++ | +++ | ||
| Flerårige vandrende arter: | |||||||
| Overjordiske stengelutløpere | ++(+) | 0 | 0 | ++ | +++ | ||
| Underjordiske stengelutløpere: | |||||||
| Følsomme** | + | 0 | 0 | + | +++ | ||
| Motstandsdyktige** | +++ | +++ | ++(+) | +++ | ++(+) | ||
| Rotutløpere | +++ | +++ | ++(+) | ++(+) | +(+) | ||
* Her tenkes spesielt på balderbrå, ** Mht. jordarbeiding
Gradering av mulighetene til utvikling av formeringsdyktige ugrasplanter:
+++ = størst muligheter, ++ = litt begrensende muligheter, + = noe muligheter, 0 = ingen muligheter
Se også tekstboks 2.9
Sommer- og vinterettårige ugras
For alle vekstskifteomløp vil de vinterettårige ugrasene stort sett være vanskeligere å holde tilbake enn de sommerettårige. Dette skyldes naturligvis at disse ugrasene kan spire og etablere seg gjennom hele vekstsesongen.
De sommerettårige artene etablerer seg hovedsakelig i kulturvekster som også dyrkes som sommerettårige, dvs. poteter og grønnsaker, og vårsådde korn og oljevekster. I disse kulturene vil også vårspirte vinterettårige arter etablere seg. Ved dyrking av høstsådde vekster som korn, oljevekster eller høstsådde fang- og grønngjødslingsvekster, vil de vinterettårige artene være dominerende. Jordarbeiding, men også kraftig regnvær om høsten stimulerer disse artene til spiring. Vinterettårige arter kan også dukke opp i eldre eng, men spiller liten rolle. Sommerettårige ugras som smånesle, svartsøtvier og stivdylle stresses hardt av kornet utover vekstsesongen, men i radkulturer trives de godt.
Som nevnt tidligere, er en del arter svært tilpasningsdyktige, meldestokk er et godt eksempel. Andre eksempler er vassarve, som er skyggetolerant, og klengemaure som er vel tilpasset rikelig nitrogen.
Toårige ugras
Myrtistel, vegtistel og krusetistel er alle toårige tistelarter. Myrtistel (myr og vassjuk jord) og vegtistel (tørrere steder) finnes oftest i beitemark og naturlig eng. Krusetistel er mest vanlig i yngre eng og i åker med «dårlig» jordarbeiding. Balderbrå er nok tradisjonelt den mest kjente representanten blant de toårige artene. Den er her i landet et typisk ugras i høstkorn og første års eng. I åker der jorda blir grundig bearbeidd, spiller den ingen rolle i vårsådde kulturer, noe den ville ha gjort, om den var vinterettårig, slik enkelte hevder. Hvis man derimot ikke foretar jordarbeiding om høsten og arbeider jorda lite om våren, kan overlevende balderbrå bli plagsom også i vårsådde kulturer. I andre land, gjerne lenger sør, er det ikke uvanlig at balderbrå setter frø i vårsådde vekster. Der får balderbrå følgelig mer preg av å være en vinterettårig art.
Flerårige ugras i eng
Av flerårige arter, vil stedbundne arter (bl.a. løvetann, høymole, groblad), og vandrende arter med overjordiske stengelutløpere (bl.a. krypsoleie), og arter med underjordiske stengelutløpere som er følsomme for jordarbeiding (bl.a. stornesle, ryllik, stormaure), hovedsakelig finnes i enga. Noen karaktertrekk ved ugras som vokser i eng:
Stedbundne ugras formerer seg seg hovedsakelig med frø og trenger ro i minst to år før frøproduksjonen kommer i gang. Denne tilstanden finner de i enga, men sjelden i åker. Redusert jordarbeiding og direktesåing gjør imidlertid at problemene med slike ugras i åker øker.
Vandrende ugras med overjordiske stengler (krypsoleie): Stolonene lever ikke lenger enn en vegetasjonsperiode, og nye planter utvikles fra nodiene.
Vandrende ugras med underjordiske stengler (rhizomer): Stornesle, nyseryllik og stormaure er følsomme for jordarbeiding. De kan finne sin plass der hvor det er lite jordarbeiding, for eksempel eng. Andre arter er motstandsdyktige mot jordarbeiding: For eksempel har kveke stort potensiale for oppformering etter jordarbeiding, og kan derfor ofte finnes i ettårige kulturer.Hvorfor mange flerårige arter trives så dårlig der hvor det er åpen åker kan være vanskelig å skjønne. Noe av forklaringen kan være at disse artene tåler nedmolding dårligere enn nedkutting (ved høsting av eng).
Flerårige ugras i åker
De mest problematiske flerårige artene, dvs. arter med underjordiske stengler som er motstandsdyktige mot jordarbeiding (kveke, hestehov), og arter med formeringsrøtter (åkertistel, åkerdylle), kan en vente å finne i nær sagt alle kulturvekster. Flerårig, vekstkraftig eng kan riktignok virke sanerende på flerårige arter som åkertistel, åkerdylle og kveke.
Når planter etablerer seg for første gang under uforstyrrede forhold, vil den ene arten avløse eller utkonkurrere den andre – det en kaller suksesjon. Til en viss grad, avhengig av hvilke kulturvekster vi snakker om, finner vi bruddbiter av slike suksesjoner i agroøkosystemene. Denne rangordningen, det at ulike planter kommer til ulike tidspunkt i en tidsrekke, og hvordan ugrasartenes egenskaper henger sammen med forholdene på vokseplassen, blir nærmere beskrevet i de følgende avsnittene. Ugrasproblemene på en gård kan langt på vei gi informasjon om hvordan kulturvekstene blir dyrket.
En åker som får stå urørt, vil bli invadert av planter som etter hvert vil dekke åkeren. Dette skyldes dels frøspredning fra omgivelsene, dels framspiring fra frø i åkerjorda (frøbanken) eller fra vegetative formeringsorganer til flerårige ugras. Ugras representerer ofte de tidligste stadiene i en slik gjengroing av en åker. Denne reetableringen av vegetasjon på en forstyrret jord (for eksempel etter jordarbeiding) kalles sekundær suksesjon. Primær suksesjon skjer bare dersom det i utgangspunktet ikke finnes noen rester av vegetasjon på stedet fra før (jfr. pionerplantene etter istida). De ulike tiltakene vi setter inn mot ugras (for eksempel harving) kan anses som hindringer for videre sekundær suksesjon. Hurtigvoksende, kortlivede planter, som ettårige og noen flerårige arter, forekommer hyppigere i starten på suksesjonen enn i senere stadier. I flerårig eng for slått eller beite forandrer både kulturen og ugrasvegetasjonen seg gradvis etter som tida går. Dersom vegetasjonen får fritt spillerom uten noen form for inngrep, vil gras- og busksamfunn overta, for til slutt å ende i et stabilt skogsamfunn (se figur 2.14). Suksesjonsforløpet kan i hovedsak forklares som resultat av konkurranse mellom plantene i plantesamfunnet. Planter som får et forsprang i vekst og utvikling ved å spire tidlig (for eksempel meldestokk), har fordel av dette i konkurranse med andre planter som spirer senere. I et lukket vegetasjonsdekke, for eksempel en tett eng, er det således ofte vanskelig for planter som spirer opp sent fra frø, å etablere seg på grunn av konkurranse fra den etablerte vegetasjonen. Derimot kan vi i eng ha innslag av flerårige arter som løvetann og høymole som utvikler seg i takt med enga. Disse artene kan imidlertid også komme til senere, og deretter stadig øke i omfang.
Generelt har alle planter tilpasset seg sine spesielle vokseplasser, basert på at tilgjengelige ressurser, som energi, karbon, vann og mineralnæringsstoffer, er blitt utnyttet på en best mulig måte for planten. Dette gir seg utslag i at de ulike planteartene og populasjonene får en spesiell oppbygning (morfologi), blomstringsfrekvens, regelmessighet i frøspiringen (periodisitet) og fysiologisk tilpasning. Disse livsstrategiene er knyttet til en bestemt genetisk utrustning (DNA) som gir dem økologiske likhetstrekk. Kunnskap om plantenes primærøkologiske strategier gir oss en nøkkel til å forstå strukturen og dynamikken i ulike plantesamfunn og økosystemer, agroøkosystemer inkludert. Faktorene forstyrrelse (for eksempel ulike mekaniske tiltak i en åker, beiting, plantesjukdom, tråkk, forurensning, vind, erosjon og brann) og stress (ytre begrensninger som hemmer tørrstoffproduksjonen i alle deler av vegetasjonen, som lav temperatur, skygge, vannmangel og næringsmangel) bestemmer i stor grad hvilke arter som vokser hvor. Kombinasjonsmulighetene mellom disse to faktorene utgjør tre forskjellige hovedklasser av strategier for plantene (tabell 2.2):
a Konkurransesterk strategi
b Stresstolerant strategi
c Ruderal strategi
Tabell 2.2 Inndeling av livsstrategier hos planter i etablert fase
| Intensitet av forstyrrelse | Intensitet av stress | |
|---|---|---|
| Lav | Høy | |
| Lav | A) Konkurransesterk strategi (eks. stornesle) |
B) Stresstolerant strategi (eks. tyttebær) |
| Høy | C) Ruderal strategi (eks. groblad der det er mye tråkk) | D) Ingen levedyktig strategi (årsak til mangel på karplanter i Antarktis) |
Se også tekstboks 2.10 Mer om livsstrategier hos planter: K–S–R–modellen
I 1974 lanserte den engelske økologen J.P. Grime K-S-R–modellen, der bokstavene står for plantenes mulige strategier: Konkurransesterk, Stresstolerant og Ruderal strategi. «Ruderal» har noe med rydning å gjøre, steder Lids flora kaller «skrotemark», der naturlig vegetasjon er sterkt forstyrret eller ødelagt av inngrep, for eksempel på tomter, fyllinger og avfallsplasser. Ytre faktorer som kan påvirke en vegetasjon deler Grime inn i to hovedkategorier: stress og forstyrrelse. Med stress menes varige (eller permanente) ytre begrensninger som hemmer plantenes biomasseproduksjon i alle deler av vegetasjonen, for eksempel lav temperatur, skygge, vannmangel og næringsmangel. I eksempelvis en fattig myr er det hele tida næringsmangel. I en tørr furuskog er det ofte både næringsmangel og vannmangel. Ved forstyrrelse blir hele eller deler av plantebiomassen ødelagt som følge av plantespisende dyr (herbivorer), plantesjukdommer, tråkk, forurensning, vind, frost, uttørking, erosjon eller brann. Harving og pløying blir i denne sammenheng regnet som en forstyrrelse. Det samme gjelder slått ved grasdyrking. Som tabell 2.2 viser, kan en ha ulike intensiteter av stress og forstyrrelse i etablert fase, dvs. når plantene har kommet over spirestadiet. Ugraset vil falle i gruppe A eller C, eller kombinasjon av A og C. Bare tre av kombinasjonene gir levedyktige vokseforhold for plantevekst. Der både alvorlig stress og forstyrrelse forekommer samtidig, er ingen vegetasjon mulig.
Kombinasjonene A, B og C i tabell 2.2 kan framstilles som henholdsvis hjørnene K, S og R i et trekantdiagram (se NO FILENAME). Hvert hjørne av trekanten representerer ekstremer av utviklingsspesialiseringene. Ifølge K-S-R-modellen vil en vegetasjon utvikle seg på et bestemt sted til en gitt tid, som resultat av en likevekt mellom intensitet av stress (varig begrensning av produksjonen), forstyrrelse (fysisk ødeleggelse av vegetasjonen) og konkurranse (naboene kaprer de samme ressursene). Over et lengre tidsrom skjer det en endring av artssammensetningen og de genetiske egenskapene til vegetasjonen ved selektiv utdøing og/eller innvandring av ulike plantearter. Der hvor det er lite stress og forstyrrelse, vil planter med gode konkurranseegenskaper invadere vokseplassen, og utviklingen vil gå i retning av monokultur. Levedyktig strategi til en viss planteart kan plasseres i en gitt posisjon inni trekanten i forhold til koordinatene i trekanten. I den posisjonen er det en form for likevekt mellom konkurranse, stress og forstyrrelse for vedkommende art.
I den virkelige verden varierer K-S-R-likevekten fra plass til plass, selv innen ett og samme plantesamfunn, både gjennom døgnet og gjennom årstider. Et plantesamfunn kan bestå av plantearter med svært ulike strategier. Ofte ligger det inne en «forsinkelse» i vegetasjonsendringen i forhold til stress og forstyrrelse.
Konkurranse. Produktive, uforstyrrede vokseplasser (for eksempel i nærheten av gjødselhauger) vil bli kolonisert av robuste, flerårige planter med høy potensiell veksthastighet (som stornesle). Sonene både over og under jorda blir okkupert av en tett, hurtigvoksende biomasse. Det er ofte høy dødelighet av planteindivider som blir fortrengt av naboene. De utdøende individene er gjerne de som er følsomme for patogener eller plantesjukdommer. Det kan også forekomme utsulting av rotsystemdeler, på grunn av lokal næringsmangel. Det vil skje en utvelgelse mot genotyper med stor morfologisk fleksibilitet. Overvintring skjer gjerne ved hvilende knopper og frø, mens andel årlig produksjon viet til frø er liten. Ved mangel på ressurser, som for eksempel næringstoffer, blir det en hurtig nedgang i utviklingen av blad og røtter.
Stress. I markert kontrast til produktive vokseplasser er vokseplasser hvor alvorlig stress begrenser plantevekst, som for eksempel kronisk mangel på nitrogen og/eller fosfor, eller tørke. Bare lave «kapringshastigheter» for de begrensede ressursene er mulig for stresstolerante plantearter (som bergknapparter og grasarten finnskjegg). Både overlevelse og reproduksjon er avhengig av plantenes kapasitet til å forbli levende gjennom lange perioder med liten vekst. Hos slike planter blir gjerne vekst «koplet» fra ressursinntaket. Vern mot beiting av plantemasse er også typisk. Stresstolerante planter overlever vinteren med røtter og ofte vintergrønne blad. Ved mangel på ressurser blir det liten respons på utviklingen av blad og røtter.
Forstyrrelse på vokseplassen favoriserer genotyper der hurtig vekst og tidlig reproduksjon øker sannsynligheten for at tilstrekkelig avkom produseres for overlevelse og reetablering av populasjonen. Vokseplasser som i utgangspunktet er næringsrike, men med hyppige forstyrrelser (som for eksempel på jordbruksarealer, hager, stier med mye tråkk, og/eller der tidsbegrenset tørke kan forekomme), koloniseres altså av arter med ruderal strategi. Blant disse finner vi også efemere plantearter, dvs. arter som overlever tørketid i form av frø (mange av våre ettårige ugras), knoller eller løker (f.eks. strandløk), men som ellers ikke har noen spesiell tørketilpasning. Andel årlig produksjon viet til frø er stor, og plantene overlever vinteren i form av hvilende frø. Mangel på ressurser gir hurtig innskrenking av vegetativ vekst med påfølgende blomstring og frøsetting.
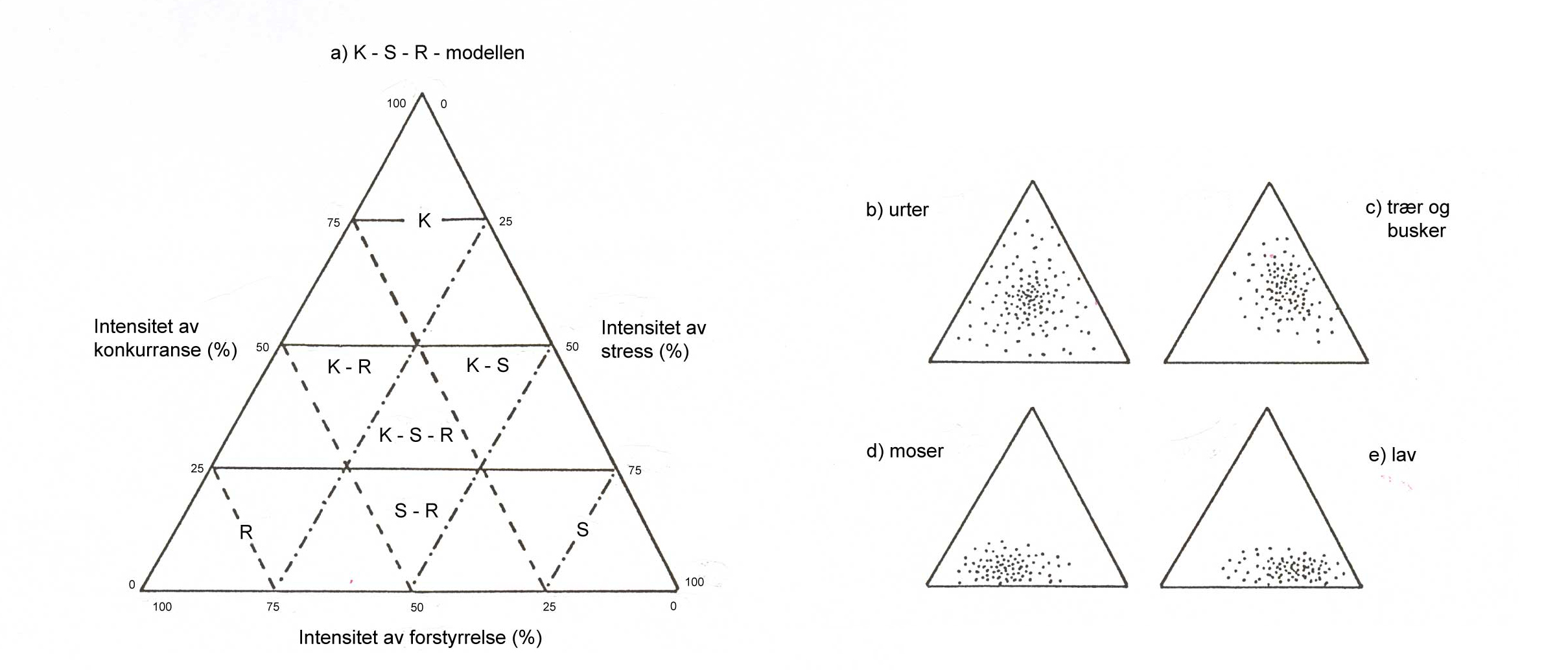
Summen av levedyktige frø av en planteart i jorda eller på jordoverflaten kalles frøbanken av vedkommende art. «Innskuddet» i denne banken er produksjonen av frø fra plantene som vokser på stedet, og frø som kommer inn utenfra, for eksempel gjennom husdyrgjødsel, uren såvare og frø som blåser inn. Ved studier av hvordan arter utvikler seg i en bestand et bestemt år eller over flere år, kan frøbanken gi et uttrykk for endringene som har skjedd i den aktuelle tidsperioden. Størrelsen av frøbanken og hvilke arter som forekommer, gjenspeiler tidligere års og dagens drift av et dyrkingsareal.
Frøformering er den eneste formeringsmåten for omkring 90% av ugrasartene. Flerårig vandrende ugras har i tillegg vegetativ spredning, eksempelvis rotutløpere. Følgelig er hovedkilden til nyspirte ugrasplanter frø spredd på jordoverflaten og/eller frø tidligere nedgravd i bakken. Blomsterplantene blant ugraset utmerker seg gjerne ved en stor frøproduksjon. Mens kornplantene normalt produserer omkring 50 frø per plante, må vi hos de fleste ugras regne med flere tusen, ti tusen eller hundre tusen frø per plante. Korsmo har i boka Ugras i nåtidens jordbruk (1954) tall for frøproduksjon hos de fleste ugras. Som eksempler kan nevnes løvetann 3000, vassarve 15 000 og groblad 21 500 frø per plante i gjennomsnitt for planter som vokser sammen med kulturvekstene, og opp til 700 000 frø på spesielt store planter av burot. Det kan også være stor variasjon innenfor en art, for eksempel kan frøproduksjonen til gjetertaske variere mellom 2000 og 40 000 frø per plante. Det er særlig i år med åpen åker at jorda blir tilført store mengder nye ugrasfrø.
De fleste frø drysser av plantene på selve vokseplassen, gjerne før kulturen er høstet. Jorda kan også få tilførsel ved ulike former for spredning, som for eksempel ved vind (løvetann), dyr/menneske (då-arter), gjødsel (høymole), såvare (floghavre) og landbruksmaskiner (floghavre). De fleste ugras er svært seiglivete og lunefulle med hensyn til framspiring. Dette gjelder særlig floghavre. Regulering/bekjemping av dette ugraset er til og med nedfelt i egen forskrift: Forskrift om floghavre.
Å hindre at frøene i det hele tatt blir tilført jorda, vil følgelig være et viktig tiltak for å holde størrelsen på frøbanken nede. Effektiv ugrasregulering i selve såbedet før frøsetting, og å hindre frøtilførsel utenfra, er eksempler på slike tiltak. Når vi diskuterer frøbank, vil spiringsbiologien til ugrasfrøene stå sentralt. Men her må det også legges til at spiringsbiologien til ugrasfrøene ikke er tilstrekkelig utforsket. Frøet hos de fleste ugras er mer eller mindre spiretregt (dormant), og spirer lite eller ikke straks etter at det er modent.
At levende frø ikke spirer med en gang, kan skyldes egenskaper ved frøet (primær frøhvile) eller egenskaper ved miljøet omkring (sekundær frøhvile).
Primær frøhvile
Denne hvilen utvikles i frøet under modningsprosessen og hindrer at frøet spirer, selv om miljøet omkring er optimalt. Årsakene til primær frøhvile kan være flere (figur 2.16):
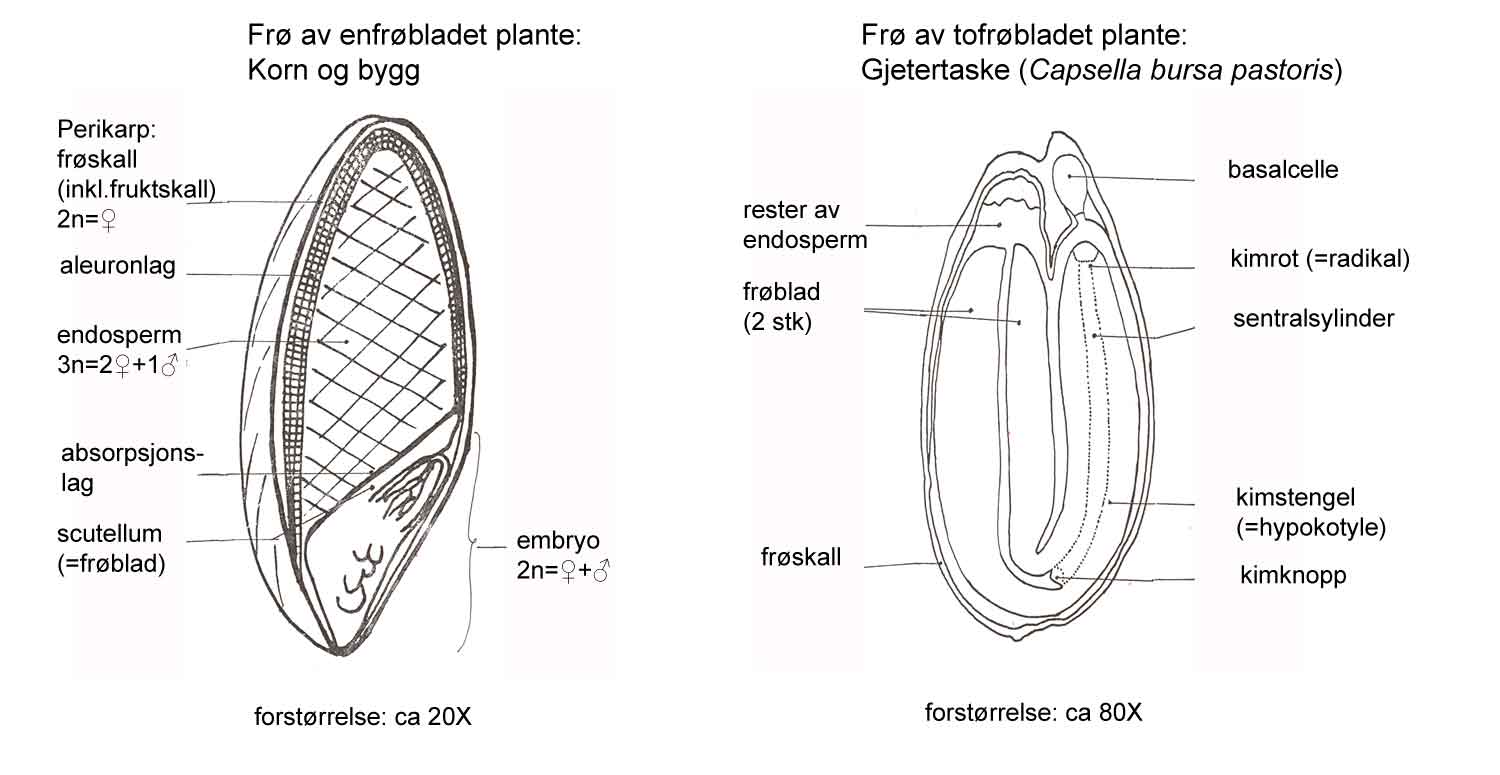
Sekundær frøhvile
Betingelsene for frøspiring er ikke til stede i miljøet omkring. Dette kan skyldes viktige faktorer som mangel på fuktighet og lys, feil lyskvalitet, for lavt oksygennivå, for lav temperatur eller hemmende (allelopatiske) stoffer i jorda. (figur 2.17).
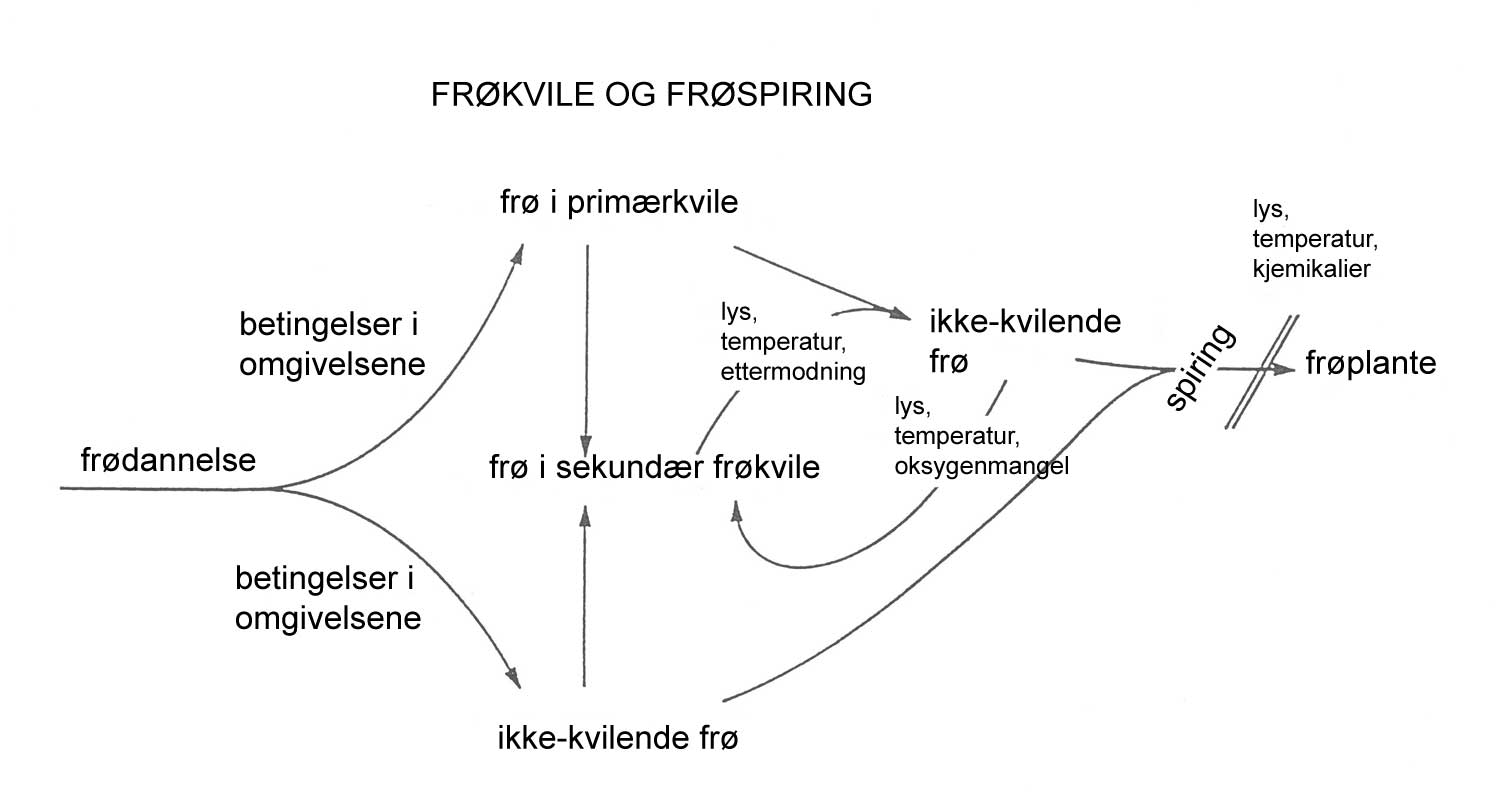
Figuren viser overgangen fra ?frø i primærhvile? eller ?ikke-hvilende frø? til enten spiring eller til ?frø i sekundær frøhvile?. For frø i sekundær frøhvile? kan hvilen bli brutt vha. endringer i miljøet.
Karakteristisk for frøhvilen hos ugrasfrøet er ellers at den kan variere sterkt, ikke bare mellom arter, men også innen samme art. Selv frø fra samme plante, eller i alle fall fra samme populasjon, kan spire til ulike tidspunkter. Noen frø spirer straks etter modning, mens en større del spirer først tidligst neste vår etter overvintring. Det er svært vanlig hos ville planter at frøene etter spredning må gjennom en periode med lav temperatur (stratifisering) før de kan spire. Under naturlige forhold vil frøene for disse artene da hindres i å spire om høsten. Endelig kan en del frø bli liggende i jorda i flere år og kanskje ikke spire før tredje, fjerde året eller senere, selv om frøet ligger både passe dypt og ellers har gode spirevilkår.
Hos de fleste ugras er det en sterk periodisitet i springen: Sommerettårige ugras har for eksempel spiretopp tidlig vår, mens vinterettårige ugras har en mindre topp tidlig vår og en større topp sen sommer (figur 2.18). Denne periodisiteten i spiringen tyder på at frøet, på grunn av arvelige forskjeller, må få ulike ytre impulser i kortere eller lengre tid, før spiringen tar til. Temperatursvingningene og daglengdeskiftene mellom årstidene har trolig mest å si.
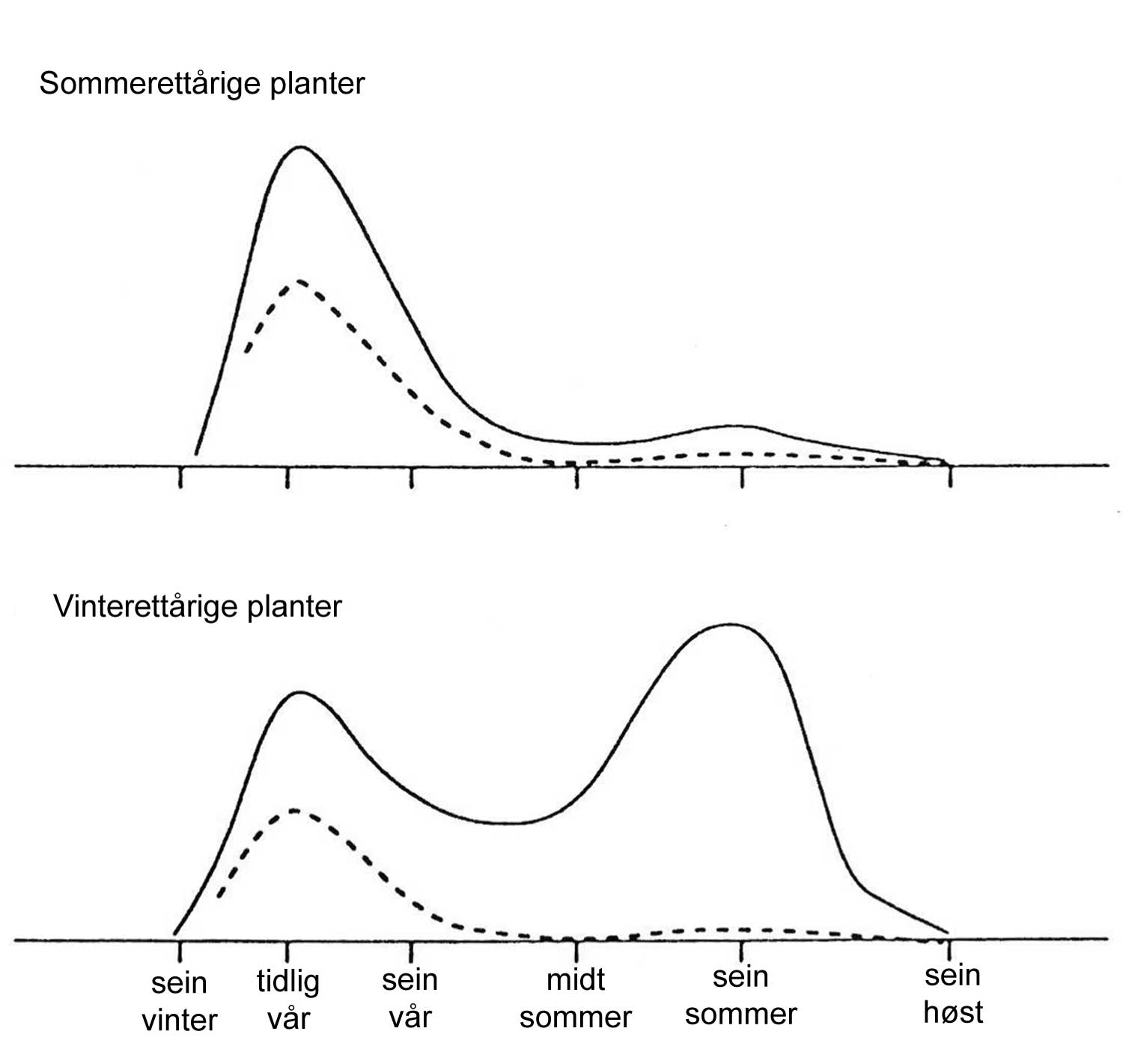
At det finnes ulike mekanismer som hindrer frø i å spire, har en viktig økologisk funksjon: Disse mekanismene, både de som ligger i selve frøet og i miljøet omkring, hindrer at en stor del eller alt frø, spirer på en gang. Hvis dette skjedde, og forholdene senere ikke var til stede for vekst, ville arten bli totalt utryddet på vokseplassen. Denne strategien er spesielt viktig for ettårige arter, siden disse må satse alt på ett kort, nemlig frøformering, i kontrast til flerårige arter, som også kan overleve ved flerårige røtter og jordstengler. For både ettårige og flerårige arter representerer frø den eneste «linken» mellom generasjonene, der utveksling av arvemateriale foregår.
Frøenes levealder
Det er en av naturens mekanismer at en levende organisme som et frø, kan være spireklart i lang tid, for så å spire når forholdene ligger til rette for det. Et imponerende eksempel er frø av arktisk lupin (Lupinus arcticus S. Wats) hvor det er funnet 10.000 år gamle frø som fremdeles var levedyktige. Også frø av ugras kan bli svært gamle, bl.a. er det funnet spiredyktige, 1700 år gamle frø av meldestokk og linbendel ved arkeologiske utgravninger. Som tabell 2.3 viser, er det stor variasjon mellom lagringsevne til ulike ugras.
Tabell 2.3 Prosent levedyktige frø etter ulik lagringstid ved nedgraving i jord
(middel av 2 og 15 cm dybde) i Fairbanks, Alaska (etter Conn og Deck 1995)
| Art | Systematisk navn | Prosent levedyktig frø | |||||
| 0 år | 2,7 år | 3,7 år | 4,7 år | 6,7 år | 9,7 år | ||
| Floghavre | Avena fatua | 97 | 4 | <1 | <1 | <1 | 0 |
| Gjetertaske | Capsella bursa-pastoris | 87 | 46 | 43 | 30 | 18 | 2 |
| Kvassdå | Galeopsis tetrahit | 46 | 0 | 0 | 0 | 0 | 0 |
| Kveke | Elymus repens | 27 | <1 | 0 | 0 | 0 | 0 |
| Linbendel | Spergula arvensis | 94 | 35 | 30 | 29 | 18 | <1 |
| Meldestokk | Chenopodium album | 100 | 49 | 25 | 35 | 17 | 4 |
| Tunbalderbrå | Chamomilla suaveolens | 85 | 60 | 61 | 25 | 20 | 1 |
| Tungras | Polygonum aviculare | 61 | 13 | 9 | 7 | 3 | 1 |
| Vassarve | Stellaria media | 89 | 62 | 39 | 26 | 26 | 5 |
| Vindeslirekne | Polygonum convolvulus | 69 | 6 | 5 | 5 | 3 | 1 |
Undersøkelsen viser en stor nedgang i levedyktighet, fra 6.7 til 9.7 lagringsår for noen av artene. Selv etter 10 år i nedgravd tilstand, var 13 av de undersøkte 17 artene fremdeles i stand til «å videreføre slekten». Selv om tallene for floghavre er lave, er det viktig å være klar over at frø av denne arten kan leve ganske lenge når de blir pløyd ned. Ei floghavreplante kan produsere opp til 500 frø.
Frøspiring og miljøfaktorer
Det første som skjer når frø «våkner til liv», er at frøet tar opp vann. Dernest vil frø bare spire innenfor et visst temperaturintervall, som da gjerne er tilpasset det klimaet vedkommende art hører hjemme i. Forholdet til temperatur blir ofte delt inn i minimum-, optimum- og maksimumstemperaturer. Også døgnvariasjoner i temperatur vil påvirke frøspiringen. For eksempel er det vist at åkertistelfrø spirer raskere hvis jordtemperaturen varierer mellom dag og natt.
En tredje viktig faktor er lys. Både lysmengde og -kvalitet (forholdet mellom rødt og mørkerødt lys) vil påvirke spireprosessen. Ikke alle ville arter reagerer likt på lys. En undersøkelse har vist at for 75 % av viltvoksende ettårige og flerårige arter ble spiringen stimulert ved lystilgang, 22,5 % var upåvirket og 2,5 % viste spirehemming ved lystilførsel. Frøene til de fleste artene som reagerer på lys, har oftest to kjennetegn: A) De er viltvoksende og B) Frøene er små. Hvis slike frø spirer for dypt nede i jorda, vil de dø fordi opplagsnæringen (som ofte er fett) i frøet blir oppbrukt før spirene når jordoverflaten. Lyskravet for å starte spireprosessen vil derfor sikre at de bare spirer når de er i nærheten av jordoverflaten. At en del frø bare spirer når temperaturen i jorda varierer mye mellom dag og natt, er også med på å sikre at disse artene bare spirer når de er i nærheten av overflaten. Lenger ned i jorda vil temperaturvariasjonen mellom dag og natt være mye mindre. Frøene til de fleste kulturplanter har ikke noe lyskrav, fordi vi gjennom foredling har selektert bort denne egenskapen. Frø kan også reagere på ulik lyskvalitet. Blant annet vil mye langbølget, mørkerødt lys hindre en del frø i å spire. Den økologiske konsekvensen av at frø reagerer på denne måten, er at mye mørkerødt lys «forteller» frøet at det allerede er en del planter som har etablert seg på vokseplassen. Altså er signalet til frøet at «nå er det ikke noe fordelaktig å spire, sjansen for å bli utkonkurrert av andre planter er for stor».
Også andre faktorer som for eksempel tilgang på oksygen, og spirehemmende stoff i jorda (allelopati) kan påvirke frøspiringen.
I radkulturer er en form for ugrasregulering å dekke til jorda med enten døde planterester («mulching») eller en underkultur (levende plantedekke). Slike former for jorddekke vil påvirke mange forhold i jorda, bl.a. lys og temperaturforhold.
Tabell 2.4 viser resultatene fra en amerikansk undersøkelse av hvilken effekt dødt og levende dekke av lodnevikke hadde på ugrasveksten. Kort oppsummert kan resultatene forklares slik: A) Et levende plantedekke reduserer sollyset mest, i tillegg til at også forholdet mellom rødt og mørkerødt lys endres mest der, i retning mørkerødt (forholdet minker i tallverdi). B) For både dødt og levende dekke er temperatureffekten noenlunde den samme: Maksimums- og minimumstemperatur henholdsvis synker og stiger (nærmer seg hverandre), noe som gjør at temperaturvariasjonen gjennom døgnet blir mindre. C) Dødt dekke vil redusere spiring og vekst hos ugraset, men i mindre grad enn et levende dekke. De ulike effektene på ugraset skyldes ikke bare virkning på frøspiringen, de ulike formene for dekke vil også i stor grad påvirke veksten til ugraset etter oppspiring. For eksempel vil mange av de ugrasfrøene som spirer i et levende dekke dø på grunn av lysmangel.
Tabell 2.4 Effekt av ulike typer plantedekke (død og levende lodnevikke), sammenlignet med bar jord,
lys- og temperaturforholdene ved jordoverflaten og ugrasvekst (etter Teasdale og Daughtry 1993)
| Klimafaktor / ugrasvekst | Bar jord | Lodnevikke | ||
| Dødt dekke | Levende dekke | |||
| Stråling (sollys) | 100 % | 17 % | 0,60 % | |
| Lyskvalitet (Rødt / Mørkerødt) | Lite påvirket | Redusert | ||
| Jordtemperatur: | Maksimum | Redusert | Redusert | |
| Minimum | Svak økning | Svak økning | ||
| Forskjell gjennom døgn | Redusert | Redusert | ||
| Effekt på ugraset | «Ingen» | Middels | Sterk | |
Spiredyp
Det optimale spiredypet for ugrasfrøet er jamt over svært grunt, det ligger for de fleste arter mellom 0 og 1 cm. Mange av våre vanligste åkerugras spirer for eksempel ikke om frøet ligger dypere enn 2–3 cm. Som tabell 2.5 viser, vil noen arter, for eksempel då-artene, spire også fra enda dypere sjikt.
Tabell 2.5 Ulike ugrasarters maksimale spiredyp rangert i spiredypklasser
| Dyp | Art | Frøspiring: Maksimum dyp (cm) | |
| Norsk navn | Systematisk navn | ||
| Små | Burot* | Artemisia vulgaris | På overflaten |
| Stivdylle | Sonchus asper | 0-1,0 2 | |
| Tunbalderbrå | Chamomilla suaveolens | 0,5 2 | |
| Hestehov* | Tussilago farfara | 1,0 2 | |
| Haremat | Lapsana communis | 1,0 2 | |
| Ryllik* | Achillea millefolium | 0-2,0 2 | |
| Åkersvineblom | Senecio vulgaris | «på overflaten eller små dyp» 1 | |
| Gjetertaske | Capsella bursa pastoris | 2,0 1 / «små dyp» 2 | |
| Pengeurt | Thlaspi arvense | 2,0 1 / 1,0 2 | |
| Rødtvetann | Lamium purpureum | 2,0 2 | |
| Åkerminneblom | Myosotis arvensis | 2,0 2 | |
| Åkersennep | Sinapsis arvensis | 2,0 1 | |
| Middels | Balderbrå | Matricaria maritima | 2,0 1 / 4,0 2 |
| Vanlig hønsegras | Polygonum persicaria | «små dyp» 2 | |
| Engsoleie* | Ranunculus acris | «små dyp» 2 | |
| Åkerdylle* | Sonchus arvensis | 0,5-3,0 2 | |
| Løvetann* | Taraxacum officinale | 2,0-3,0 2 | |
| Linbendel | Spergula arvensis | 3,0 2 | |
| Kvassdå | Galeopsis tetrahit | 1,0-4,0 2 | |
| Vassarve | Stellaria media | 3,0 2 | |
| Groblad* | Plantago major | 4,0 2 | |
| Krypsoleie* | Ranunculus repens | 4,0 2 | |
| Større | Småsyre* | Rumex acetocella | 5,0 2 |
| Meldestokk | Chenopodium album | 5,0 1 og 2 | |
| Åkertistel* | Cirsum arvense | 6,0 1 / 1,0 2 | |
| Klengemaure | Galium aparine | 6,0-9,0 2 | |
| Dåarter spp. | Galeopsis spp. | ||
| Tungras | Polygonum aviculare | ||
| Vindelslirekne | Polygonum convolvulus | ||
| Kveke* | Elymus repens | ||
| Høymole* | Rumex longifolius | ||
| Floghavre | Avena fatua | 17,5 1 / 25,0 2 | |
1Kilde: Radosevich og Holt (1984), 2Kilde: Korsmo (1935), *Flerårige arter
Dersom vi hindrer ny frøsetting og spredning utenfra, minker altså tilførselen til frøbanken. I tillegg kan en del ugrasfrø følge med avlingen og bli ødelagt på forskjellig vis, for eksempel ved ensilering. Dessuten kan jorda tappes for frø ved at de spirer, og deretter ødelegges ved ulik jordarbeiding, eller rett og slett fortæres av jordboende organismer. Helbrakk, som er «ekstremversjonen» av jordarbeiding, der ingen kulturvekst finnes og jordarbeidinga stadig gjentas gjennom hele vektsesongen, er selvfølgelig det mest effektive tiltaket for å redusere frøbanken. I en undersøkelse ble det vist at ett år med helbrakk reduserte frøbanken med om lag 50 %. Helbrakk er imidlertid en gammeldags, lite miljøvennlig og økonomisk ulønnsom metode. I et engelsk såkalt «sylinderforsøk» ble det funnet en årlig reduksjon i frøbanken på 32 % i «forstyrret jord», med en halveringstid på 2 år (den tida det tar for at frøbanken skal bli halvparten så stor). I «uforstyrret» jord derimot, ble den årlige reduksjonen bare 12 %, og halveringstida hele 6 år.
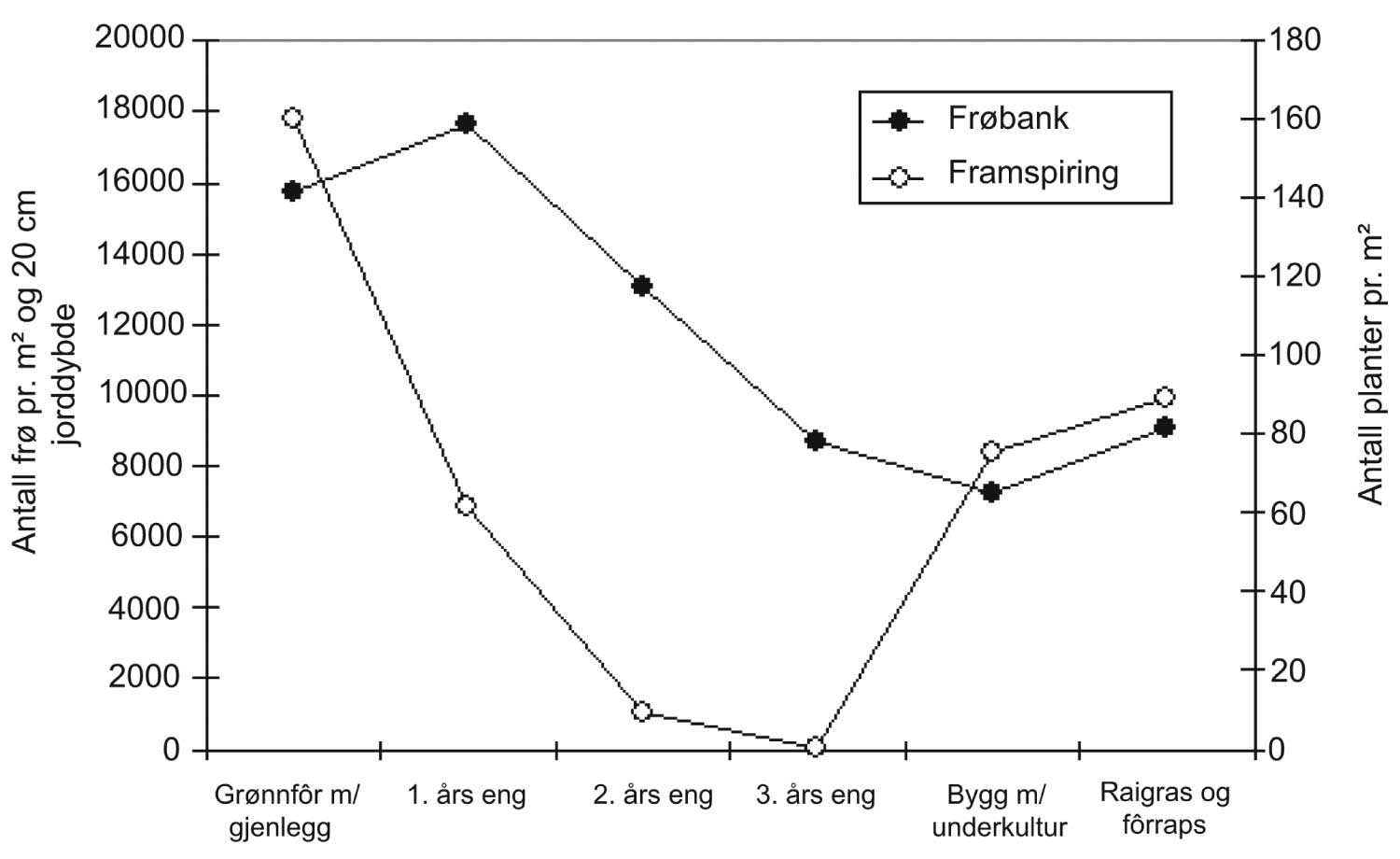
(etter Sjursen 2001)
Selv om hovedmengda av frø stammer fra tidligere år, vil utviklingen over tid endre seg i takt med de ulike kulturene i omløpet. I et økologisk drevet areal ved Universitetet for miljø- og biovitenskap (tidligere Norges landbrukshøgskole) ble det etter en engperiode på tre år med minkende frøbank, hovedsakelig bestående av ettårige ugrasarter, funnet en rask økning i neste periode med korn og ulike radkulturer (figur 2.19). Hvor mye frøbanken økte i åkerkulturene var selvfølgelig avhengig av hvor god kontroll en hadde på ugraset i disse kulturene. Maksimum ble i gjennomsnitt oppnådd første året etter åpenåkerperioden, altså i 1. års eng, og minimum første året etter engperioden, i bygg med underkultur. (Tidspunktene for maksimum og minimum er forskjøvet ett år i forhold til de reelle, kort og godt fordi analysene ble gjort først tidlig om våren året etter at frøene var produsert) Da var frøbanken redusert til ca. 40 % av maksimumsnivået. Legg også merke til svingningene i antall framspirte ugras, som er resultatet av jordarbeidinga og konkurransen som ettårig ugras møter i ulike kulturer i omløpet. I 3. års eng, hvor jorda ikke ble arbeidet og hvor det var god vekst i de konkurransesterke, flerårige kulturplantene, ble det f.eks. minimal framspiring av ettårige ugras, og tilsvarende mindre bidrag til frøbanken. I grønnfôr med gjenlegg, som er en relativt lysåpen og konkurransesvak kultur, ble det maksimal framspiring av ettårige ugras, med tilsvarende bidrag til frøbanken, noe som altså kunne registreres i etterfølgende kultur.
Utviklingsstadium og variasjon i næringsinnhold
Når nye planter utvikler seg fra frø, tappes frøet for næring. Summen av næring i frø og plante reduseres også inntil et bestemt stadium. Deretter vil næringsinnholdet øke. Dette stadiet, kompensasjonspunktet, inntreffer omkring frøbladstadiet (figur 2.20). Plantene er da svært følsomme for forstyrrelser, for eksempel jordarbeiding.
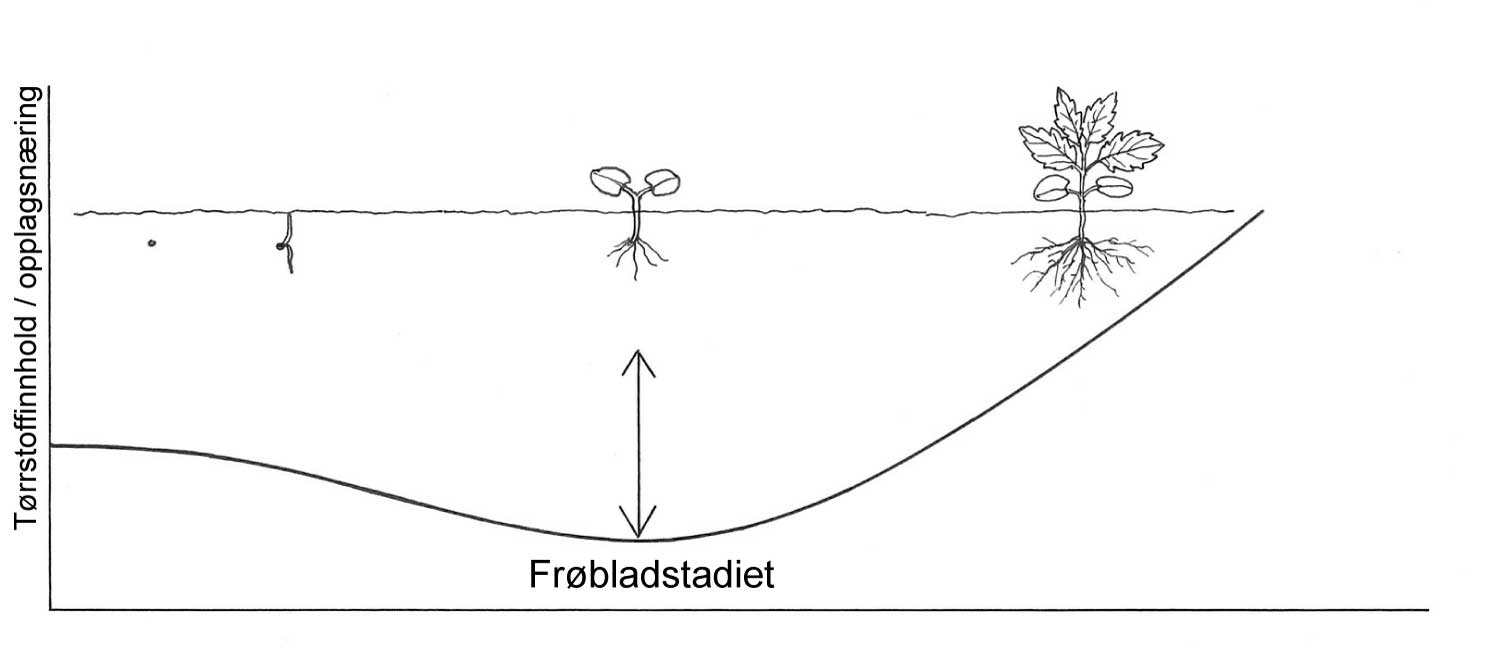
Levealder
Et frø kan spire bare en gang. Dersom den unge planten blir hindret i å utvikle seg, har ikke frøet flere sjanser til å gi opphav til en ny plante. En knopp på en jordstengel, rot eller knoll, kan heller ikke bryte mer enn en gang, men skulle dette skuddet bli ødelagt, finnes det gjerne en ny knopp på den samme organdelen som kan bryte i steden. Et vegetativt formeringsorgan er således ikke uten videre ferdig med livet fordi om en av knoppene blir borte. I forhold til de fleste frø er levealderen til vegetative formeringsorganer vesentlig kortere. Levealderen synes å variere en del, avhengig av både plantearten og de aktuelle jordarbeidingstiltakene og vokseforholdene. I svenske forsøk er det for eksempel funnet at korte jordstengler hos kveke dør etter at de nye lysskuddene var etablert, dvs. etter ca. 2 år. Lengre jordstengelbiter derimot kunne være i god stand også ved utgangen av dette andre året. I forsøk med 5 cm lange røtter av åkerdylle og åkertistel, og jordstengler av hestehov med 5 knopper, er det funnet at disse formeringsorganene bare i få tilfeller ble eldre enn 2 år.
Tidsaspekt 1: Utviklingsstadium og variasjon i næringsinnhold
Når nye planter utvikler seg fra et vegetativt formeringsorgan blir dette, på samme måte som et frø, tappet for næring. Mens frøet etterpå går helt til grunne, kan altså det vegetative formeringsorganet samle ny næring fra den overjordiske, grønne delen av planten og leve videre for kortere eller lengre tid. Nye formeringsorganer vokser også fram, men egne lysskudd makter disse ikke å sette før etter at morplanten har nådd en viss størrelse.
Både tørrstoffmengde og næringsinnhold i vegetative formeringsorganer varierer med utvikling av planten. Fra å ligge på et middels nivå før de nye skuddene begynner å vokse, passerer mengda av tørrstoff og næring et minimum en eller annen gang i vekstsesongen. Når lysskuddene blir store nok til å sende mer næring (assimilater) ned i røttene eller jordstenglene enn disse organene bruker opp, stiger tørrstoffmengda igjen. Plantene tåler minst å bli forstyrret i veksten når de vegetative formeringsorganene er mest tappet for opplagsnæring.
For de fleste plantearter har en ikke noe eksakt svar på hvor store plantene er når minimumspunktet for tørrstoffinnholdet passeres. Noen viktige ugras er undersøkt mer enn andre. Svenske forsøk har vist at kveke passerer dette stadiet når lysskuddene er 12–15 cm høye og har 3–4 blad. Tilsvarende minimum for åkerdylle og åkertistel vil du finne i figur 2.21. Undersøkelser har vist at hestehov har tilsvarende minimum når det største bladet er mellom 4 og 10 cm bredt, og åkersvinerot når bladrosetten har 6–12 blad. Et praktisk problem er at de mest følsomme stadier ofte ikke kommer før etter normale tider for etablering av vårsådde vekster. Da kan det være vanskelig å sette inn nødvendige tiltak.
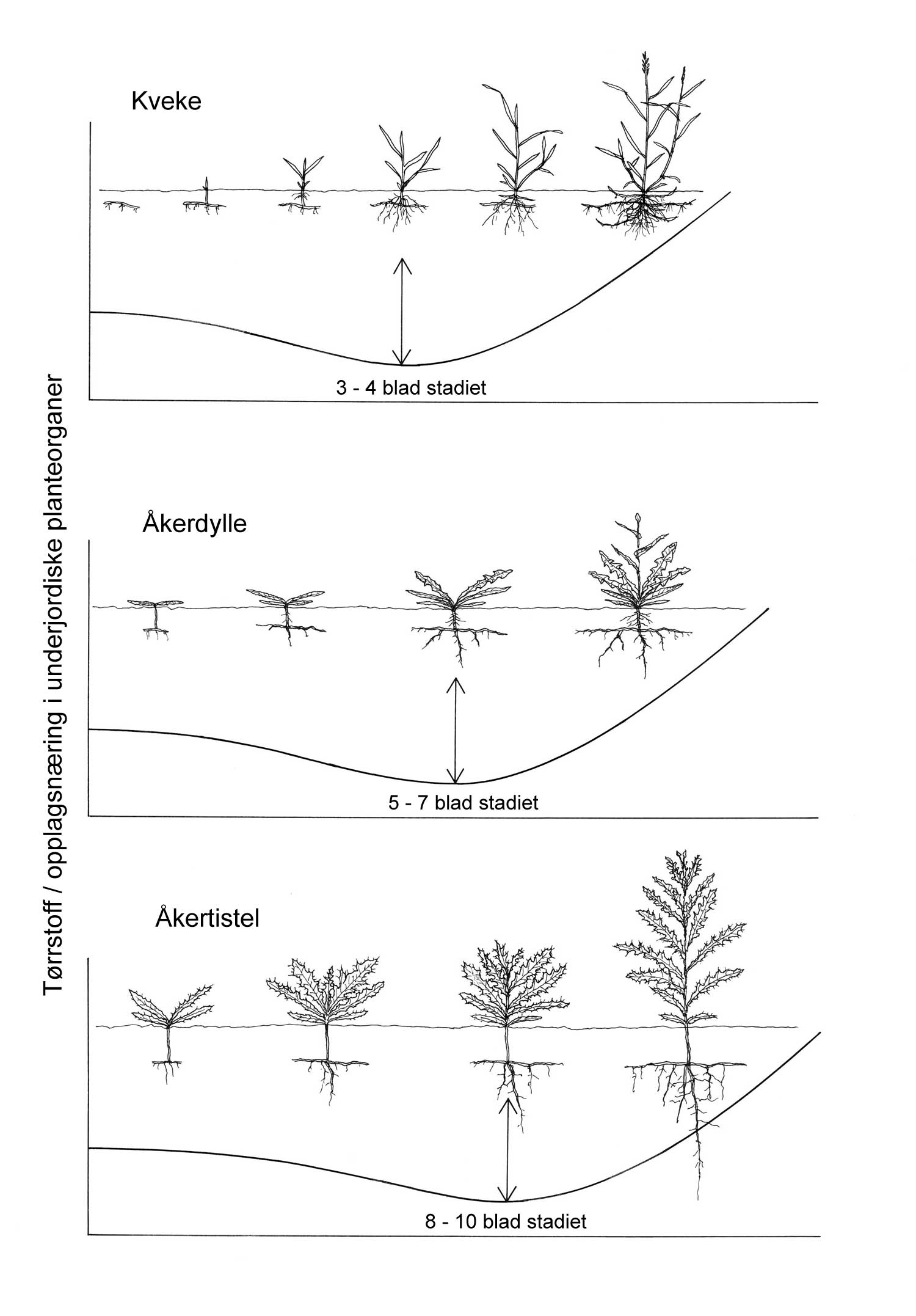
Hvordan kunnskapen om kompensasjonspunktet for en art kan utnyttes i praksis, er vist for kveke i figur 3.26 (stubbarbeiding om høsten). Figur 2.26. viser effekten av gjentatt jordarbeiding ved ulike utviklingstrinn (fra 1–2- bladstadiet til 6–7-bladstadiet) for kveke. Hvis jordarbeidinga gjøres for sent ser vi at tørrstoffmengda i underjordiske plantedeler øker. Tilsvarende figurer kunne man også lage for andre arter med krypende formeringsorgan, men effekten vil bli forskjellig for de ulike arter. Den relativt gruntvoksende åkerdylla kan for eksempel utarmes med færre behandlinger om disse utføres tidlig i vekstsesongen. Mer dyptvoksende arter som åkertistel, hestehov og åkersnelle utarmes saktere enn kveke.
Av hensynet til andre faktorer er det sannsynligvis lite aktuelt med helbrakk i økologisk dyrking, men figur 2.26 viser at en gjentatt jordarbeiding i deler av vekstsesongen kan ha mye for seg. Ved flere gangers jordarbeiding bør den første sørge for sterk oppdeling slik at mange knopper aktiveres, noe som fører til stort forbruk av opplagsnæring. I kombinasjon med en etterfølgende konkurransesterk vekst, kan svært god effekt på ugraset forventes.
Generelt vil nye jordstengler eller formeringsrøtter til flerårige ugras for alvor begynne å vokse først etter at kompensasjonspunktet er passert. Før den tid har altså plantene bare den utarmete mor-jordstengelen eller mor-roten å lite på. Det er ikke sikkert at alle flerårige planter har akkurat dette mønsteret, men for de plantene vi vet noe om, er dette nyttig kunnskap. Ved å utføre jordarbeiding, radrensing, luking og/eller hakking når plantene er svakest, oppnår vi størst virkning med minst innsats.
Tidsaspekt 2: Knopphvile/dormans
På samme måte som frø, kan også de vegetative formeringsorganer være i mer eller mindre hviletilstand. De fleste arter er mest spirevillige om våren og tidlig sommer. Utover høsten kan det utvikles knopphvile (dormans) med forskjellig styrke, som bl.a. er artsavhengig. Kveka har for eksempel lite dormante knopper, selv sent på året. Dette benytter vi oss av i høstbrakkingen ved at vi kutter jordstenglene (rhizomene) med harv eller fres for at så mange knopper som mulig skal bryte. Når lysskuddene er blitt passelig store (opp til 3–4 blad), ødelegger vi dem ved ny jordarbeiding. Studier i Norge har vist at åkertistel og åkerdylle har likhetstrekk når det gjelder evne til å sette nye skudd fra røtter (figur 2.22). Figuren viser at disse artene har stor evne til å sette skudd tidlig om våren men at evna taper seg raskt utover høsten (egentlig en dropp i mai–juni pga. de første skuddene, men evna stiger igjen og når en topp i juli–august før den da raskt faller igjen). Et generelt råd for disse to artene er at mekaniske tiltak bør settes inn fra våren og framover til omkring begynnelsen av august. Etter denne tid inntrer i alle fall for åkerdylla en utpreget skuddhvile. Dette betyr i praksis at uansett hvor mye røttene til åkerdylla kuttes opp, så vil det ikke komme opp lysskudd på denne årstida. Dette vil først skje neste vår. Forsøk har vist at hestehov har tilsvarende årsrytme som åkertistel. Siden åkerdylle blir mer eller mindre spiretreg utover sensommeren, kan vi derfor ikke vente at oppdeling av dyllerøttene om høsten, selv med ny jordarbeiding senere, skal gi færre planter året etter, slik tilfellet er med kveke. Hvis vi ikke pløyer rotbitene dypt ned, slik at de nye lysskuddene får lang vei opp til overflaten, kan resultatet heller bli omvendt.
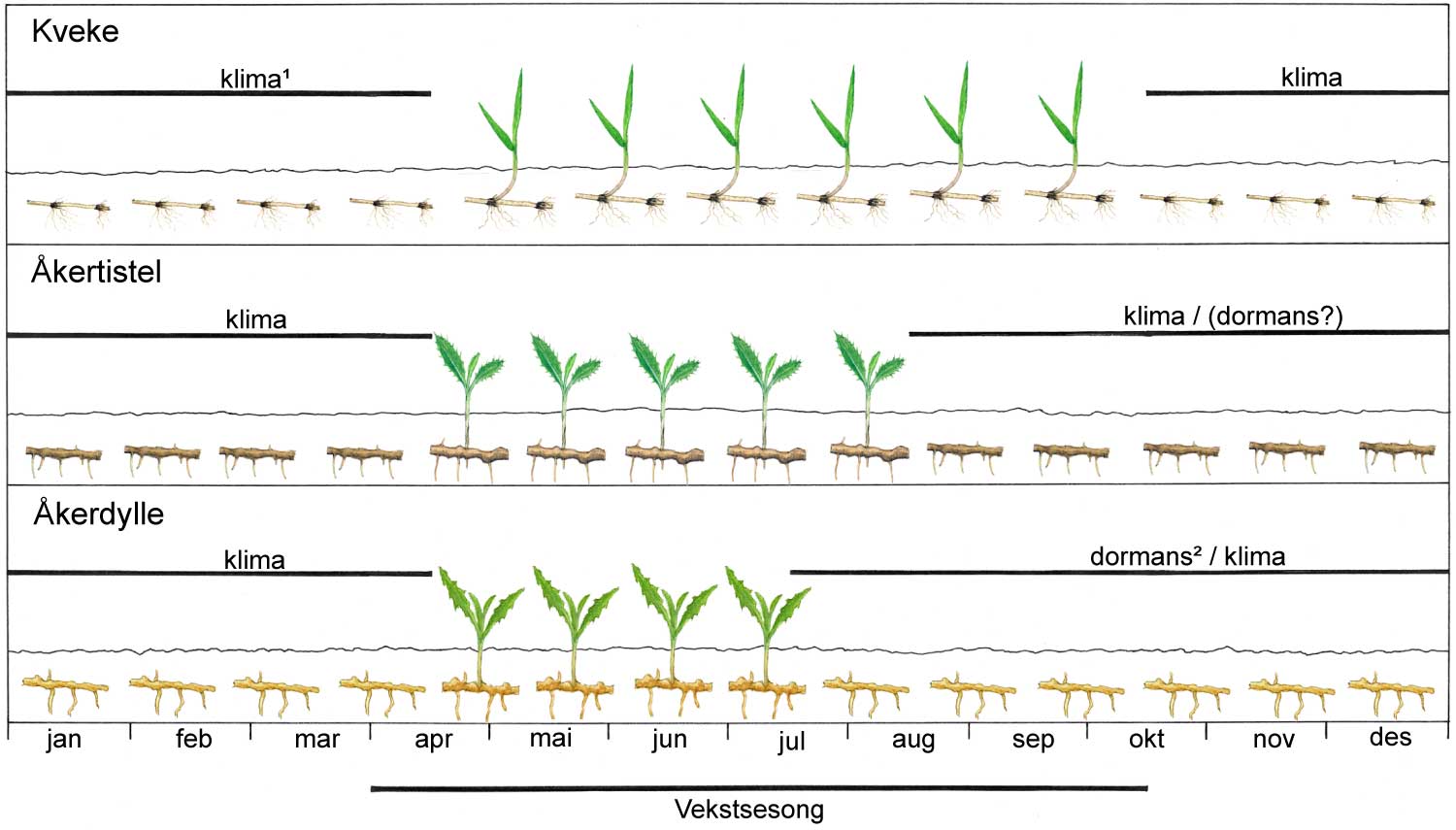
Hvor dypt formeringsorganet ligger
Som figur 2.23 viser, går rotsystemet hos åkertistelen betydelig dypere enn hos åkerdylle og kveke. Det er funnet vertikale røtter av åkertistel helt nede på 2–3 meters dyp i tyngre jordarter. Motstykket til dette er kveka som har hele massen av jordstengler i pløyedybde. De horisontale formeringsrøttene hos åkertistel finnes hovedsakelig 15–30 cm nede i jorda. At disse røttene ligger så dypt, betyr at de mest vanlige jordarbeidingsredskaper ikke når ned og får kuttet dem opp. Formeringsrøttene til åkerdylle ligger som jordstenglene hos kveke adskillig grunnere, i all hovedsak i de øverste 10–15 cm. Englodnegras, storkvein og vasslirekne er andre eksempel på arter med relativt gruntliggende formeringsorgan, mens de hos hestehov og åkersnelle som hos åkertistel går betydelig dypere.
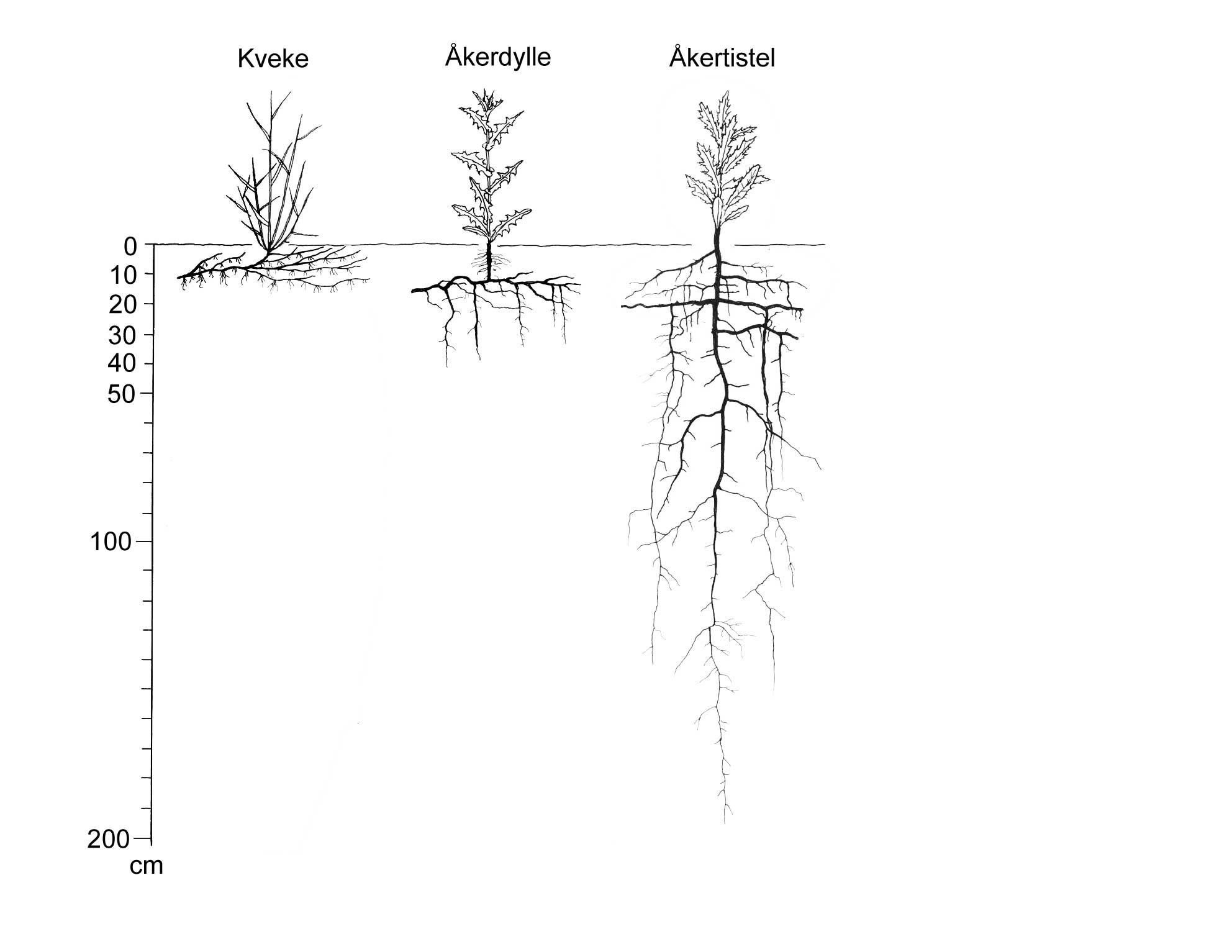
Oppkutting av vegetative formeringsorganer
De svake periodene for ugraset som er omtalt her, kommer altså fram ved at knoppene på de vegetative formeringsorganene bryter og utvikler lysskudd. Jo kortere jordstengel- eller rotbitene er, jo mindre næring er det bak hvert skudd, og jo mindre tåler planten å bli forstyrret. Ved å kutte opp jordstenglene og røttene i mindre biter, vil flere knopper bryte. Oppdelingen av formeringsorganene vil altså øke effekten av de etterfølgende tiltakene, så sant de blir satt inn til rett tid. Gjør vi ikke det, vil oppdelingen av jordstenglene/ røttene tvert om føre til flere planter.
Undersøkelser viser at åkerdylle både har flere skudd per cm rot og spirer hyppigere fra større dyp, enn tilfelle er for åkertistel (se figur 2.24). Om man skal anbefale en sterkest eller svakest mulig oppdeling av røtter eller jordstengler, avhenger av hva som skjer etterpå. Hvis man ønsker å sulte ut ugraset mest mulig, er det fordelaktig med sterk oppdeling av røttene, og etter siste behandling dyp nedpløying av rotbitene. På den annen side vil en sterk oppdeling av røtter med påfølgende etablering av en kulturplante hvor man ikke kan sette inn tiltak, for eksempel korn, være uheldig. Effekten av ulik oppdelingsgrad og nedgravingsdybde for kveke er vist i figur 2.25.
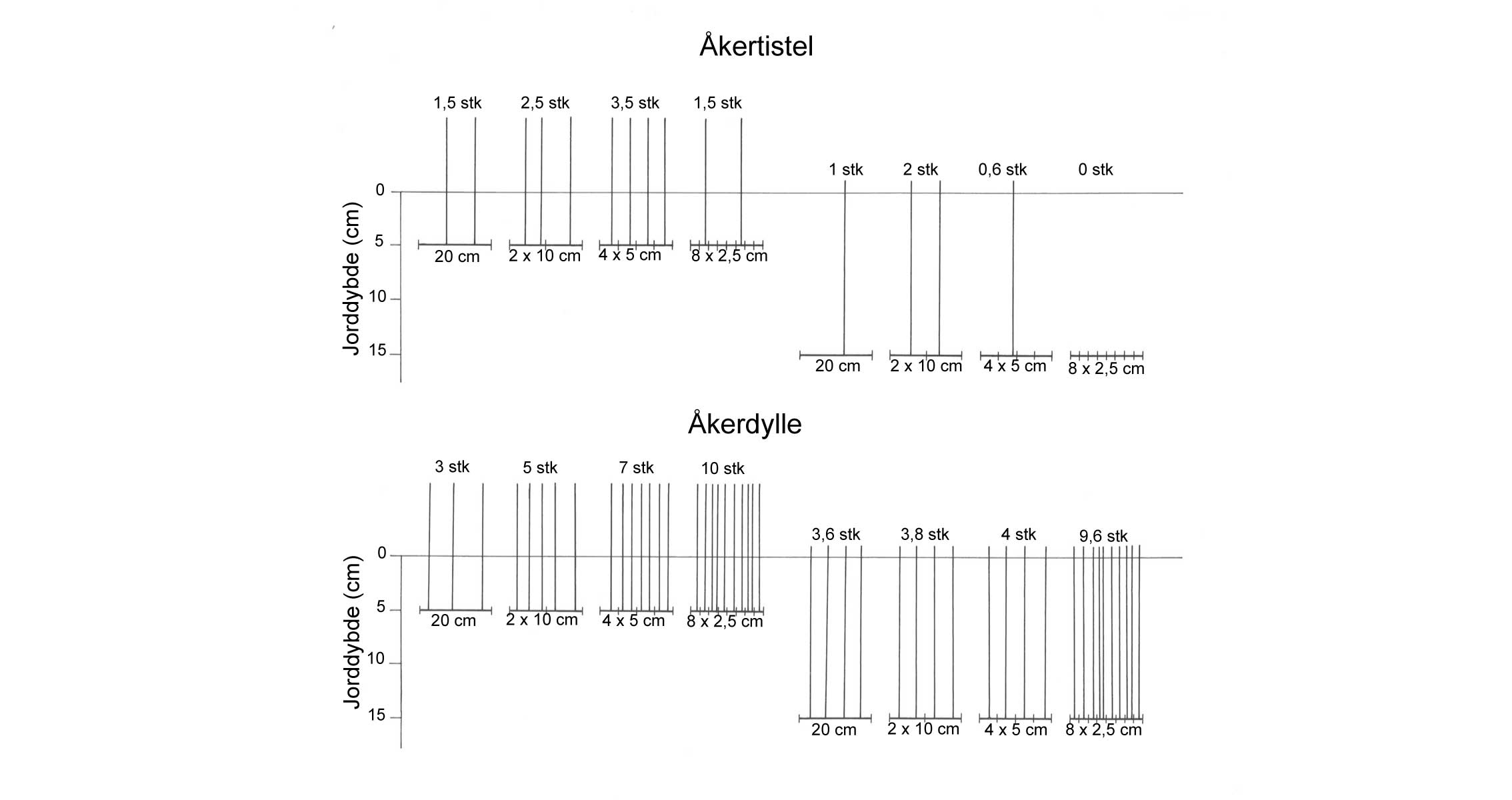
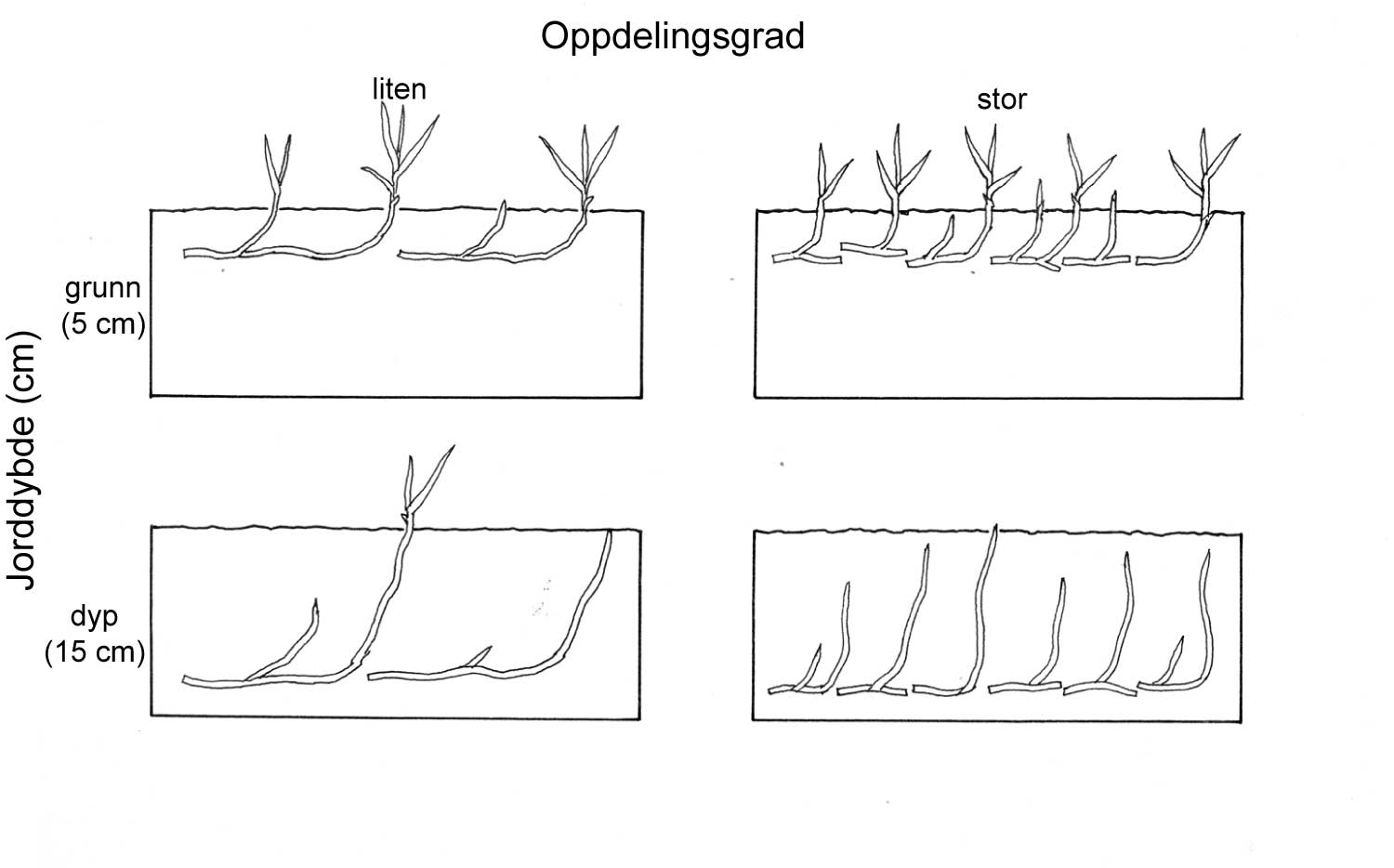
Hvor store dyp et vegetativt formeringsorgan greier å sende opp skudd fra, varierer med plantearten og størrelsen på organdelen. Jo større den er, jo dypere kan den ligge, og enda klare å sette nye lysskudd. Felles for alle ser likevel ut til å være at dersom de kommer for dypt til å sette lysskudd første året, hjelper det ikke om de kommer opp på et lageligere dyp året etter. Da er de døde. Vegetative formeringsorganer skiller seg derfor sterkt fra de fleste frø, som kan ligge mange år i jorda uten å ta skade.
Gjentatt jordarbeiding for «å sulte ut» ugraset
Som figur 2.26 viser, kan man utarme kveka ganske raskt hvis man for eksempel bruker våren og forsommeren til jordarbeiding på en optimal måte. Hvis man gjennomfører helbrakk (en metode som på grunn av kostnadene og miljøeffektene ikke er særlig aktuell i dag) vil det meste av kveka være borte i løpet av en vekstsesong. For andre ugrasarter vet vi mindre, men den gruntvoksende åkerdylla utarmes nok raskere, dvs. ved færre behandlinger, enn for eksempel arter som åkertistel, åkersnelle og hestehov med dypere rotsystem. Figuren under viser at man får en raskere utsulting av kvekejordstenglene hvis man gjennomfører behandlinga ved kvekas 2 bladstadium enn ved senest 3–4 bladstadiet. Amerikanske undersøkelser har vist at åkertistelen blir raskest tappet for opplagsnæring hvis jordarbeidinga gjøres hver 3. uke.
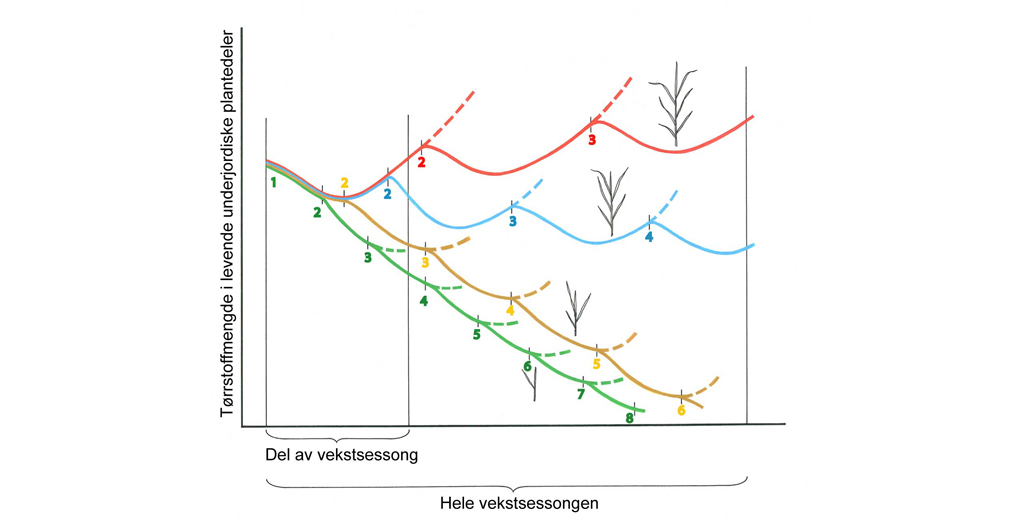
Påvirkning av klimafaktorer som temperatur og fuktighet
Kvekas vekst styres i stor grad av vær og klimaforhold. Nedre temperaturgrense for at kveka skal vokse er om lag 5 ºC. For mange andre ugras vet vi ikke de nøyaktig temperturkravene under norske forhold, men utenlandske undersøkelser kan tyde på at for eksempel åkertistel slutter å vokse tidligere, om lag 8 ºC. En annen viktig klimafaktor for vekst og utvikling er jordfuktigheten. Optimale vekstvilkår for kveka om høsten kan være både en fordel og en ulempe, sett i relasjon til bonden og ut fra «kvekas perspektiv»: Hvis det er optimale forhold for kveka utover høsten, og man ikke setter inn mottiltak, vil kveka selvfølgelig ha «gode dager» og produsere store mengder nye jordstengler og mye opplagsnæring. Hvis man iverksetter en utsultingsstrategi under optimale forhold (gjentatt jordarbeiding ved 3–4-bladstadiet), vil jordstenglene tynes svært effektivt. Hvis det derimot er veldig tørt i jorda om høsten, vil kveka vokse lite uansett, og en utsultingsstrategi vil da være mindre effektiv.
Det har vært en del fokus på muligheten til å drepe røtter og jordstengler ved uttørking på jordoverflaten, spesielt gjelder dette kveka. I Norge er ikke klimaet spesielt gunstig for en slik strategi. Klimaet vårt er ofte for fuktig. Mer informasjon om dette finnes i tekstboks 2.11.
For å kunne tørke i hjel kveka etter oppharving kreves det kveke i god vekst, tørr luft og høy temperatur. Det er derfor først og fremst vår og sommer at dette kan bety noe i kvekekampen. Ved hver harving vil bare en liten del av jordstenglene komme opp på overflaten. Vi må derfor ha lang tid med pent vær for å få et godt resultat. Det er viktig å huske på at utsatt såing for å få tid til en slik uttørking, ofte medfører redusert avling. Hvis forholdene ligger til rette for det, vil jordstengler som ligger oppå overflaten kunne drepes gjennom uttørking i løpet av noen dager. Danske undersøkelser har vist at kvekejordstengler på overflaten av tørr jord kan drepes på 4 dager i varmt vær, men at man i praksis ofte mislykkes. Forsøkene i Danmark viste videre at hvis man fikk 2 uker med tørt vær i slutten av august, hadde dette en bra effekt på jordstenglene. I september var det vanskeligere å skade jordstenglene, og i oktober var det naturligvis liten effekt. Under norske forhold kan man forvente svært vekslende resultat både mellom ulike plasser og mellom år. Forsøkene i Danmark viste at visse jordarbeidingsredskaper kan dra opp 75 % av utløperne.

I samfunn av ulike organismer vil mange arter regulere hverandre ved å produsere og frigjøre kjemiske stoffer. Dette fenomenet blir ofte klassifisert under fellesbetegnelsen kjemisk økologi. Et eksempel er insekter eller nematoder som ikke angriper visse planter pga. frastøtende stoffer (repellenter) eller toksiske kjemikalier som planten produserer. Allelopati representerer «plante- mot-plante»-aspektet på dette området. Betegnelsen allelopati ble lansert av vitenskapsmannen Molish i 1937 for å beskrive kjemisk samspill mellom planter (mikroorganismer og høyere planter), inkludert både stimulerende og hemmende effekter. I en del miljøer har uttrykket allelopati blitt brukt i snevrere forstand, begrenset til bare hemmende effekter mellom høyerestående planter. Molish sin opprinneleige og brede definisjon kan i mange tilfeller være mer hensiktsmessig å bruke, blant annet fordi mange undersøkelser har vist at allelopati direkte eller indirekte ofte involverer mikroorganismer. De aktuelle stoffene behøver ikke bare komme fra plantene, men kan også bli avgitt av mikroorganismene, som på sin side bruker plantemateriale som substrat («mat»). Det er vist at slike naturlig forekommende stoffer kan ha både en hemmende og stimulerende effekt på planter ved henholdsvis høye og lave konsentrasjoner.
Det er vanlig i litteraturen å separere samspillet mellom planter i henholdsvis konkurranse og allelopati. På friland er det svært vanskelig å skille disse to komponentene fra hverandre, men det er utviklet metoder for bruk i veksthus/laboratorier for å isolere allelopatidelen fra konkurranse.
Stoffer med allelopatisk effekt kan finnes i alle plantedeler. Om disse stoffene frigjøres til omgivelsene i tilstrekkelig mengde til å påvirke naboplantene, er fremdeles i mange tilfeller usikkert.
Stoffer med allelopatisk virkning blir ofte kalt allelokjemikalier. Allelokjemikalier blir frigjort ved fordamping, utskillelse av roteksudater, utvasking og nedbryting av planterester. Det er mye som tyder på at mange slike stoffer frigjøres sammen, og at en hemmende effekt oppstår på en additiv eller synergistisk måte. Allelokjemikaliene representerer et mangfold av ulike stoffer, fra enkle hydrokarboner til komplekse polysykliske strukturer. De representerer syrer, aldehyder, alkaloider, terpenoider og mange andre stoffer.
Det er grunn til å tro at allelopati i visse plantesamfunn er en viktig samspillfaktor, mens det i andre er av mindre betydning. Det er uenighet blant forskere om allelopatiens betydning i landbruket.
Noen godt dokumenterte eksempler:


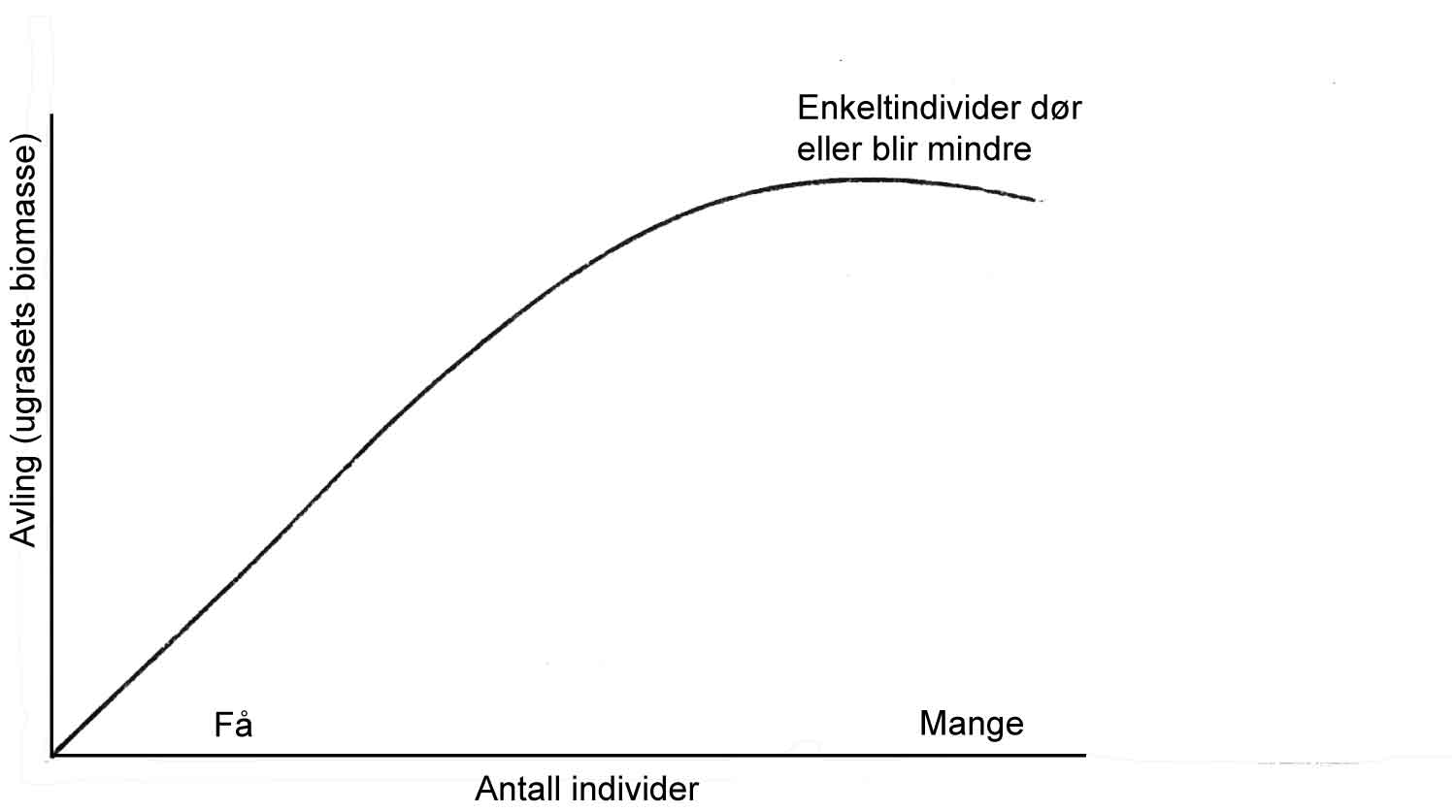
Ugraset
Den relative framspiringstida for kulturplanter og ugras varierer bl.a. med egenskapene til såvaren, såbed og dets tilberedning (eksempelvis jordart, sådybde og gjødsling) og værforhold. Skaden ugraset kan gjøre den første tida etter såing, er vanskelig å rette opp senere i vekstsesongen, selv med god gjødsling. Ved mye ugras kan gjødsling gjøre vondt verre. På den annen side kan rikelig gjødsling føre til rask og god vekst i kulturplantene, og når dette kombineres med direkte kontroll av ugraset, for eksempel harving, gi god effekt mot ugraset. Hvilke muligheter vi har for å kontrollere ugrasframveksten i de ulike kulturene, blir utførlig behandla i Bind 2–4 om henholdsvis grønnsaker og potet, korn og frukt og bær.
Mange vanlige ugras finnes i den «ruderale» gruppen, d.v.s. de som tåler forstyrrelse på vokseplassen. Artene karakteriseres gjerne med høy potensiell veksthastighet, i tillegg til produksjon av svært mange frø per plante under gunstige forhold, sammenlignet med kulturplanter (tabell 2.6). De småfrøete artene tunrapp og vassarve har for eksempel høyest maksimal veksthastighet.
Tabell 2.6 Hastighet av frøplanters tørrstoffproduksjon (ruderale arter og bygg/tomat) (etter Grime 1979)
| Arter | Frøvekt (mg) |
Maksimal veksthastighet (g per uke) |
Gjennomsnittlig relativ hastighet per uke som funksjon av bladarealet | Bladareal (g per m2 per uke) |
| Ruderale planter: | ||||
| Tunrapp | 0,26 | 2,7 | 1,7 | 120 |
| Tungras | 1,5 | 1,4 | 1,4 | 134 |
| Vanlig hønsegras | 2,1 | 1,3 | 1,3 | 177 |
| Vassarve | 0,35 | 2,4 | 2,1 | 101 |
| Kulturplanter: | ||||
| Tomat | 3,1 | 1,6 | 1,0 | 64 |
| Bygg | 37,8 | 1,6 | 0,9 | 64 |
Den nevnte effekten av gjødsling kan langt på vei forklares rent fysiologisk. For å kunne utnytte nitrogengjødsling, hovedsakelig tilført som nitrat (NO3-), må plantene ha et effektivt enzym (nitratreduktase) som kan omgjøre nitratet til en form (nitritt) som planten kan «viderebehandle». Tilførsel av nitrat øker enzymaktiviteten umiddelbart (se tabell 2.7). Dersom et meldestokkfrø havner på en næringsfattig vokseplass, vil ikke frøet utvikle seg på grunn av næringsmangel. Dette kan utnyttes ved dyrking av kulturplanter, ved å plassere gjødselen i nærheten av kulturfrøet, mens områdene omkring ikke får tilført gjødsel (jfr. bruk av kombimaskiner for såing og gjødsling ved korndyrking).
Tabell 2.7 Induksjon av nitratreduktase i planter på ruderale vokseplasser, angitt som mikromol produsert nitritt per time per gram friskvekt (etter Kinzel 1982)
| Planteart | Aktivitet av nitratreduktase | ||
| Før tilsetting av nitrat | Etter tilsetting av nitrat | Aktivitetsøkning (%) | |
| Klengemaure | 5,9 | 7,2 | 22 |
| Løvetann | 3,8 | 4,1 | 8 |
| Meldestokk | 5,3 | 6 | 13 |
| Stornesle | 12,9 | 14,6 | 13 |
| Svinemelde | 3,3 | 4,4 | 33 |
| Tungras | 5,9 | 7,3 | 24 |
I tett bestand (for eksempel en kornåker) blir det mangel på lys jo dypere ned i bestanden en kommer, avhengig av bladverkets utforming og høyde på enkeltplantene. Noen ugras (for eksempel vassarve og åkerstemorsblom) er spesialtilpasset lavere lysintensiteter enn ordinært dagslys ved å vokse optimalt ved svakere lys. Andre arter som åkerkål, meldestokk og kvassdå, øker veksten ved økende lysintensitet. Disse artene har fordeler ved tynn åker, eller når de vokser høyt i bestanden.
Et annen spesialtilpasset ugras er vindeslirekne, som kan klatre på andre planter. Ved å vokse på denne måten kan plantene redusere forbruk av karbohydrater til produksjon av styrkevev, noe som muliggjør en desto hurtigere vekst av bladverk, dannelse av blomster og deretter modne frø. I et dansk vekstskifteforsøk var vindeslirekne den eneste arten som økte biomasseproduksjonen helt proporsjonalt med antall planter uten tegn til noen «avflating» av vekstkurven (jfr. figur 2.30), hvilket gir arten en stor konkurransefordel. Dessuten har den relativt få, men store frø (1 frø = 5,0 mg), som kan spire fra større dyp (ned til 8 cm) enn småfrøete arter, for eksempel vassarve (ned til 3 cm) (se tabell 2.5 og avsnittet spiredyp).
Ugrasfloraen er i stor grad et speilbilde av driftsformen. Forekomst og betydning av ulike ugras har endret seg gjennom tida. Foruten de naturlige betingelsene, som klima og jordforhold, har både tekniske og politiske endringer i landbruket over tid påvirket ugrasfloraen.
Noen eksempler kan nevnes:
Hvilken betydning ugraset har ved økologisk drift, kan følgende talleksempler illustrere. I et økologisk felt på UMB ble det registrert bl.a forekomst av åkertistel i både omleggingsperioden og de første årene den økologiske driften var i gang. I følge tabell 2.8 fantes det ikke åkertistel da registreringene startet i 1992. Planten har etablert seg fra 1993. Fra og med 1994 var hele arealet omlagt til økologisk drift. Figur 2.31 viser samlet utvikling for alle tofrøblada rotugras. Bortsett fra en topp i 1995, synes frekvensen av slike ugras å ha stabilisert seg.
Et annet talleksempel er fra kornfelt i et kulturlandskapsprosjekt, som viser at økologisk drift byr på et større artsmangfold eller biodiversitet, enn konvensjonell drift (se figur_2.32). I åkerkantene er det klart flest arter per m² ved økologisk drift. Ved avstandene 0,5-30 m er det liten forskjell i artstetthet mellom økologisk og konvensjonell drift, dersom det er gras i omløpet. I ensidig korn er det færrest arter per m².
Tabell 2.8 Frekvens av åkertistel (antall ruter med funn av totalt 16, per år og skifte)
ved omlegging til økologisk landbruk på Frydenhaugjordet (UMB), Ås (etter Sjursen 2002)
| Skifte | Sum | ||||||
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| 1992 | - | - | - | - | - | - | 0 |
| 1993 | - | 2 | - | - | 2 | - | 4 |
| 1994 | - | 3 | 2 | - | 2 | - | 7 |
| 1995 | 1 | 4 | 1 | 1 | 2 | 1 | 10 |
| 1996 | 3 | 3 | 1 | 1 | 2 | 1 | 11 |
| 1997 | 2 | 3 | 1 | 1 | 1 | 1 | 9 |
| 1998 | 4 | 11 | - | 6 | - | 6 | 27 |
| 1999 | - | 10 | 1 | 2 | 1 | - | 13 |
| Sum | 10 | 36 | 6 | 11 | 10 | 9 | |
i korn ved ulike driftsformer (etter Sjursen 2001).
Plantesjukdommer kan forårsakes av virus, sopp og bakterier, altså de samme gruppene mikroorganismer som gir sjukdom hos mennesker og dyr. Også næringsforstyrrelser, for mye eller for lite vann og lys, for høy eller for lav temperatur, giftstoffer i naturen osv. er med på å hemme plantenes optimale utvikling. Dette skjer både direkte og indirekte ved å gjøre plantene mer mottakelige for sjukdomsangrep. Et sjukdomsangrep fører ikke alltid til tydelige symptomer. Livsfunksjoner og yteevne kan likevel være nedsatt. Motstandskraft eller resistens mot sjukdommer avgjøres av plantens gener, men også av en rekke fysiske/fysiologiske faktorer. Et menneskes motstandskraft kan bli nedsatt ved ulike former for stress. På samme måte vil noen planter lettere kunne bli utsatt for sjukdom dersom de blir stresset, for eksempel ved tørke, næringsforstyrrelser eller lignende. Kunnskap om hva som påvirker plantenes motstandskraft er viktig når vi skal forsøke å begrense sjukdomsangrepene. Alle de fine samspillene i naturen gjør at dette blir komplisert. Best mulig kunnskap om disse sammenhengene vil uansett være et viktig middel for å forebygge sjukdom hos plantene. Det vil nok være et urealistisk mål å hindre enhver form for «nedsatt helse» hos en plante, men det er et mål å strebe mot så sunne planter som mulig. I dette avsnittet vil vi særlig gå inn på livsstrategiene til de ulike sjukdomsfremkallende mikroorganismene, for å kunne forstå og dermed forebygge angrep av disse.
Soppene er organismer med stor variasjon i størrelse, form og levemåte. Blant soppene finner vi alle overganger fra encellete via mikroskopiske arter til store hattsopper. Soppene vokser på, og skaffer seg næring fra levende eller døde planter eller dyr og svært mange er viktige nedbrytere i naturen. Parasittsoppene (tekstboks 2.13) på jordbruksvekster er mikrosopper, og kan som regel bare ses som et belegg av soppvev på planta, eller vi ser bare plantas reaksjon eller symptomene på soppangrepet. De fleste parasittsoppene er samtidig patogene, dvs. sjukdomsfremkallende. Av de om lag 100 000 soppartene vi kjenner, kan noe mer enn 10 000 arter framkalle sjukdom på planter. De fleste sopper er saprofytter som lever av dødt organisk materiale, noen er parasitter uten at de dermed framkaller sjukdom. Alle sopper absorberer all sin næring fra miljøet omkring. For å ta til seg næring må soppene skille ut enzymer som bryter ned cellulose, lignin, proteiner og andre organiske stoffer til enkle sukkerarter og aminosyrer de kan ta opp. En sopp som er parasitt på en levende plante kan fortsette å vokse som saprofytt på døde planterester. Sopper på ettårige planter kan på den måten overleve fra en sesong til annen hvis de ikke danner hvilestrukturer.
En parasittsopp er en sopp som snylter på en annen levende organisme og tar næring fra denne. Noen parasittsopper lever kun på levende planter og er da biotrofe. Disse kalles obligate parasitter. Sopper som kun lever på dødt organisk materiale kalles nekrotrofe parasitter. Andre parasitter (utgjør de fleste andre sopper) kan leve på både dødt og levende organisk materiale og varierende næringsmedier. Disse kalles ikke-obligate parasitter. Av de ikke-obligate parasittene finner man sopper som lever det meste av tida på levende planter, men som kan gå over til å leve på dødt plantemateriale under spesielle omstendigheter. Disse er semibiotrofe og kalles fakultative saprofytter. Andre lever det meste av tida på dødt organisk materiale, men kan under spesielle omstendigheter angripe levende planter og bli parasitter. Disse kalles fakultative parasitter.
Mjølauke - helsefarlig «medisinplante» var tittelen på en folder utgitt i 1992 som fagnytt i plantevern, skrevet av Leif Sundheim. Teksten nedenfor er et utdrag av teksten i folderen.
Mjølauke-soppen Claviceps purpurea er en snylter på korn- og grasarter. Fra smitta blomster vokser det fram svarte hvileknoller (sklerotier) isteden for frø. Hvileknollene blir to til tre ganger så lange som frøene og stikker ut gjennom agnene i blomsten. I grasarter med små frø, slik som kvein og timotei, blir hvileknollene bare noen millimeter lange. I korn kan de derimot bli 2-3 cm lange.
Mjølauke-soppen lever i et uvanlig samspill med planter i grasfamilien. Sporene til soppen spirer på arret omtrent som pollenkorn. Lander de på et arr som er pollinert, vil de ikke kunne vokse ned i fruktknuten. Hvis sporene derimot spirer like etter at blomsten har åpnet seg, vokser spiretråden fra arret gjennom griffelen ned i fruktknuten. Etter at fruktknuten er gjennomvokst av soppen vokser det fram konidiesporer som tyter ut sammen med sukkerholdig plantesaft fra blomsten. Dette trekker til seg insekter som sprer soppen fra plante til plante og fra blomst til blomst på samme plante. Tørt vær etter blomstring fremmer denne sekundærspredningen fra smitta blomster.
Hvileknollene vokser fram fra smitta fruktknuter i takt med veksten av frøene i friske blomster. Ved frømodning stikker hvileknollene ut gjennom agnene i blomsten. De løsner lett, faller ned på bakken og overvintrer. En del hvileknoller vil følge med korn eller grasfrø i skurtreskinga, og uten skikkelig rensing vil de kunne komme ut i åkeren med såvaren. Om våren spirer hvileknoller i jordskorpa med rødfargede fruktlegemer omtrent så store som et knappenålshode (på stilk). Spiringen og sporekastingen faller sammen i tid med blomstringen av gras og korn.
Mjølaukesoppen har mange vertsplanter i grasfamilien og er lite spesialisert. Det vil si at den kan smitte fra en grasart til en annen, og fra gras til korn. Rug har krysspollinering og mer åpen blomstring enn de andre kornartene. Derfor er mjølauke mer vanlig i rug enn i bygg og hvete. Havre er sjelden utsatt for mjølauke.
I tidligere tider var mjølauke i korn et alvorlig helseproblem for mennesker. De giftige alkaloidene i mjølauke får blodårene til å trekke seg sammen slik at blodsirkulasjonen til armer og bein blir redusert. Forgiftning kunne føre til koldbrann med påfølgende tap av fingre og tær, eller skader på nervesystemet, epileptiske kramper og død. I land med mye rugdyrking var det eksempler på at tusenvis av mennesker døde som følge av mjølauke i matkornet.
Giftvirkningen på husdyr ligner de som er skildret i eldre litteratur om mennesker med mjølaukeforgiftning.
«Mutterkorn», det tyske navnet på hvileknollene, viser til bruk av mjølauke i folkemedisin i gamle dager. Uttrekk av mjølauke ble bl.a. brukt til å sette i gang fødsler og til å stoppe blødninger.
Hvileknollene inneholder flere giftige alkaloider, deriblant ergotamin og ergotolin. Disse og andre alkaloider blir i dag brukt innen medisinen, blant annet mot migrene. Selv om en plante er infisert med mjølauke, er det bare blomsten (utvikling av hvileknoller i disse) som er giftig. Soppen lever ikke i resten av planten som dermed er fri for giftstoff. Med moderne rensing av såkorn er det liten fare for å så ut hvileknoller med såkornet om våren. Derfor er mjølauke sjelden noe problem i korndyrkinga i dag. I grasfrøavlen kan det derimot fortsatt være så sterke angrep av mjølauke at det går ut over avlingene. I godt stelte beiter vil graset ikke vokse fram til frømodning, derfor får ikke soppen tid til å produsere hvileknoller. I gras på utmarksbeite vil man ofte finne noe mjølauke.
Tiltak mot mjølauke vil bli omtalt i Bind 3 som omhandler plantevern i korn.
I flere soppslekter finner vi sopparter som er i stand til å produsere giftige forbindelser, kalt mykotoksiner. Mykotoksiner i fôr og mat kan forårsake ulike plager og symptomer alt etter type og mengde. Det er også forskjeller mellom dyrearter i hvor raskt de reagerer på innhold av mykotoksiner i fôret. Reaksjoner kan være alt fra fôrvegring til abortering og problemer med lever og nyrer. Eksempler på mykotoksindannende sopp finnes i en rekke vekster over store deler av verden. Aspergillus flavus er et eksempel på en mykotoksinproduserende sopp som hovedsakelig opptrer i varmere strøk. Den kan danne aflatoksin, et svært giftig toksin som sporadisk kan finnes i f.eks. peanøtter. Ochratoksin er et annet toksin som dannes av en Penicillium-art. Dette toksinet kan blant annet forekomme i rosiner, vin og kaffe. Også korn fra våre områder kan inneholde ochratoksin, spesielt dersom kornet ikke er tørket tilstrekkelig raskt ned eller dersom det er lagret under fuktige forhold. Sopper fra slekten Penicillium vil ofte være tilstede på kornet ved høsting, men det er først og fremst på lager at enkelte av disse produserer ochratoksin. Sopper som kan skape problemer på lagret korn kalles ofte lagermuggsopper. Generelt har vi hatt lite problemer med ochratoksiner i norsk korn. På grunn av fare for dannelse av dette toksinet er det imidlertid svært viktig med rask nedtørking av kornet til 15 % vanninnnhold. De viktigste mykotoksinprodusentene i korn i Norge kommer imidlertid fra soppslekten Fusarium. Arter i slekten Fusarium angriper kornet i vekstsesongen og kan danne mykotoksiner i akset/kornet. Fusarium kalles av den grunn ofte for "åkermuggsopp". Fusarium er et stort problem i mange ulike vekster i store deler av verden.
Fusarium culmorum er en vanlig Fusarium-art under norske forhold. Denne arten danner toksinet deoksynivalenol, eller DON som det ofte forkortes til. Dette toksinet tilhører gruppa «trichothecener» som er den største gruppa Fusarium-toksiner. Arten Fusarium avenaceum er den vanligste arten på korn i Norge. Den trives godt under norske forhold, men er mindre vanlig sydover i Europa. Denne Fusarium-arten kan blant annet produsere toksinet moniliformin, et toksin som regnes for å være mindre skadelig. Generelt vil det være slik at fuktige somre fremmer angrep av Fusarium i kornet og dermed øker faren for høyere toksininnhold i kornet sammenlignet med tørrere somre. Fusarium-sporer smitter fra planterester på bakken og opp i akset ved regnsprut. Det er under kornets blomstringsperiode at kornet er spesielt mottakelig for Fusarium-infeksjon. I tillegg er det en ulempe dersom kornet blir stående lenge utover høsten i fuktig vær. At kornet er tørt ved høsting, og at fuktigere korn blir raskt tørket ned er viktig også for å unngå dannelse av Fusarium-toksiner. Fusarium stopper imidlertid å vokse/produsere toksiner ved ca 20 % vanninnhold, mens Penicillium kan danne ochratoksin helt ned imot 15 %.
Klimaforholdene i vekstsesongen er som nevnt viktige for Fusarium-infeksjon og dermed toksinnivået, men andre forhold har også betydning. Det er vist at redusert jordarbeiding, som fører til økt mengde planterester på, og i jordoverflaten, kan føre til høyere infeksjon av Fusarium og også noe høyere innhold av mykotoksiner. Vi har også sett at sprøyting med kjemiske soppmidler har ført til en økning av Fusarium-infeksjonen i kornet. Dette kan skyldes at andre sopper blir drept av soppmidlene, mens Fusarium ikke påvirkes særlig mye. Det er derfor en teori at en god del Fusarium som overlever fungicidbehandlig faktisk får mindre konkurranse om plass og næring enn den hadde før sprøyting. Vekstskifte, gjødsling, tetthet i plantebestandet, bruk av stråforkorting og ulike sortsegenskaper vil også kunne påvirke et Fusarium-angrep.
Forskning pågår fortsatt for å finne ut mer om hva som forårsaker variasjon i innholdet av Fusarium, og hvordan man kan lage prognoser for infeksjonsnivå av denne soppen.
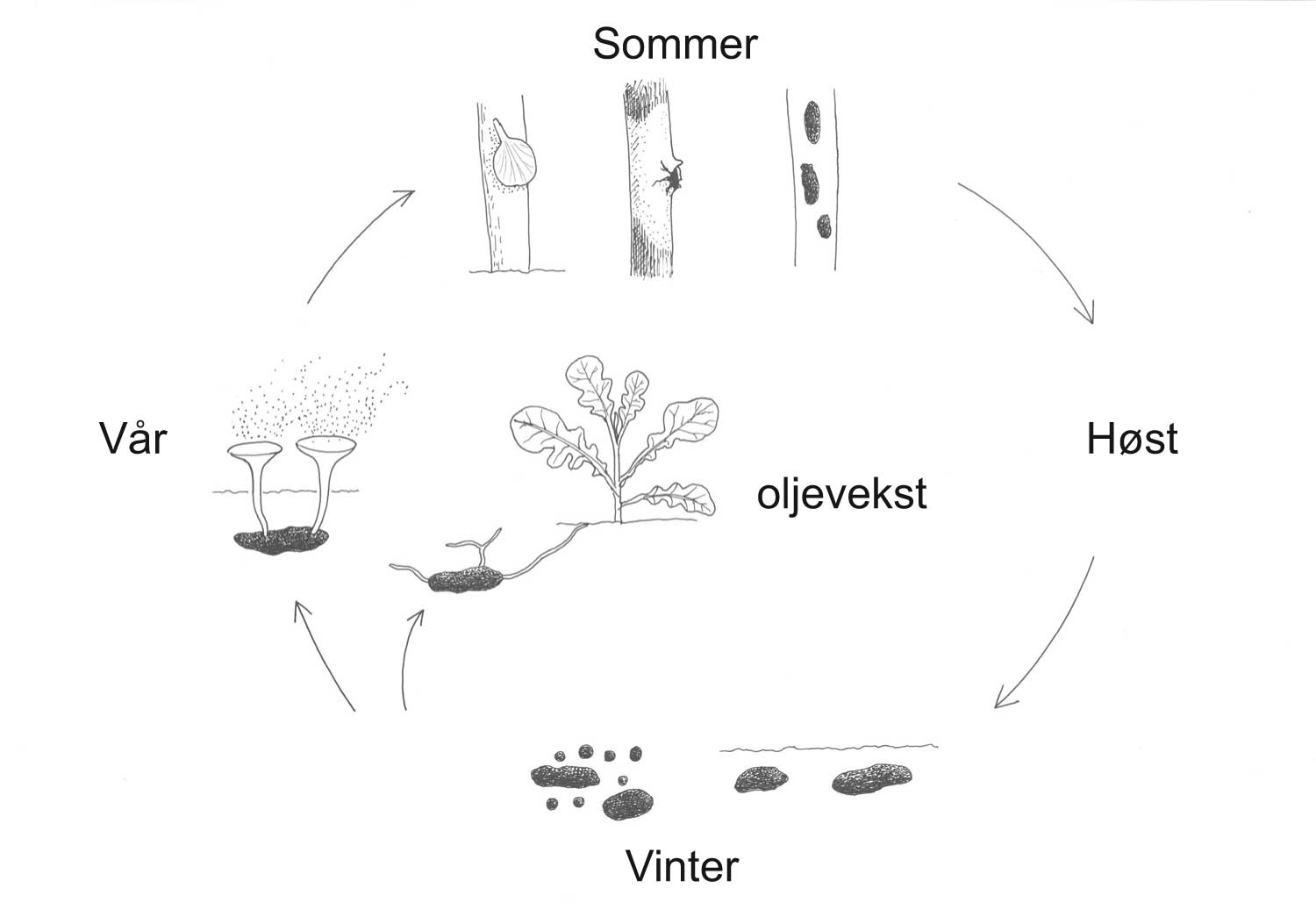
Infeksjonsprosessen
Infeksjonsprosessen innledes med at en soppspore spirer med en hyfe, og hyfespissen trenger inn i planten. Dette kan skje ved at hyfespissen trenger seg vei gjennom plantenes overhud (epidermis), eller de kommer inn gjennom naturlige åpninger i planten, for eksempel via spalteåpninger. Slike sopper kalles primære patogener. I andre tilfeller greier soppen bare å trenge inn dersom planten er såret i overhuden av maskiner, redskaper eller andre parasitter. Slike sopper kalles sekundære patogener. Ofte finner vi sekundære sopper i et plantevev som opprinnelig er angrepet av et primært patogen, noe som kan forstyrre symptombildet og gjøre det vanskelig å bestemme årsaken til infeksjonen.
Etter at planten er blitt infisert av en patogen sopp, tar det ofte noe tid før man ser resultatet i form av symptomer. Denne perioden uten synlige tegn på infeksjon, kalles inkubasjonstida. Soppen arbeider nå inne i plantevevet. Dersom soppen er aggressiv og skader eller dreper plantevevet raskt etter hvert som den trenger seg vei i planten, vil symptomene kunne bli synlige etter få dager, for eksempel som tørråte på potetblad. Inkubasjonstida er også avhengig av klimaforholdene og typen sopp eller patogen. Et eksempel på særlig lang inkubasjonstid finner vi hos stinksot, en frøoverført sjukdom som angriper høsthvete. Den kommer først til syne i akset under modning av kornplantene, selv om den opprinnelige infeksjonen startet året før under oppspiringen.
Spredning og smitteveier
De små og lette soppsporene fraktes lett med luftstrømmer og vind, fra plante til plante, åker til åker, landsdel til landsdel og land til land. Mjøldogg i bygg kan ikke leve saprofyttisk på dødt plantemateriale, derfor må den hvert år «fraktes» inn i landet med vindstrømmer fra Danmark/Sverige. Der det dyrkes vinterbygg, vil smitten kunne overleve i vinterbyggåkrene og spre seg derfra til vårbyggåkre tidlig i sesongen. Dette betyr tidligere angrep av mjøldogg i vårbyggåkre.
Vindspredning av sporer er nok den vanligste måten sopp blir spredd på, men andre veier betyr også noe. Regnsprut betyr mye for enkelte sopper som overlever i dødt plantemateriale på marka. Her blir sporene spredd ved at regndråper treffer sporulerende soppvev på marka som derved får fart og kan lande oppe på bladverket hvor soppen kan etablere seg og leve videre som parasitt. Soppsmitte fra plante til plante i samme åker blir en blanding av kontaktsmitte og spredning ved hjelp av vind eller luftstrømmer. Sopper som lever i jorda kan også angripe planter gjennom røtter. Soppslekten Fusarium inneholder flere ulike arter som er årsak til mange forskjellig sjukdommer i ulike kulturer. Den kan smitte direkte fra jord til bl.a. potet, gulrot, kløver og korn. Den samme soppen smitter også ved hjelp av regnsprut opp i kornakset. I kornet kan den overleve og spres videre dersom kornet brukes som såkorn. Infisert såkorn og såfrø er viktig som spredningsvei for en rekke ulike sjukdommer. Smitte av sjukdom kan også spres via løker og småplanter. Redskaper, maskiner, dyr og mennesker kan også være med på å spre plantesjukdommer, både over korte og lange avstander.

Sett i forhold til soppene forårsaker bakteriene færre plantesjukdommer. Noen av dem har imidlertid stor økonomisk betydning, og flere regnes som farlige skadegjørere, for eksempel i potet (ringråte). Bakterier er encellede organismer med fast cellevegg, men uten en cellekjerne. Derfor kalles de også prokaryoter som betyr «før kjerne» (rike = Prokaryotae). Formen på de plantepatogene bakteriene er stavformet. Bakterier vokser ved å dele seg i to. Noen bakterier henger sammen to og to eller flere i kjeder etter delingen. Under gunstige forhold kan bakterier dele seg en gang i timen eller oftere. Slik kan en bakterie gi opphav til noen millioner avkom i løpet av et døgn. Bakteriene danner da «kolonier», som kan ses som slimete masser med det blotte øyet. Artsgrensene er ikke alltid så tydelige, derfor har det vært mange endringer når det gjelder navnsettingen hos ulike bakteriearter. Selv om bakterier deler seg og derved har ukjønnet formering, kan også deler av arvematerialet overføres mellom en del bakteriearter, særlig blant Pseudomonas-artene.
De fleste av de rundt 1600 kjente bakterie-artene medvirker i naturens husholdning ved nyttig omsetning og nedbrytning av plante- og dyrerester, mens et hundretalls arter er sjukdomsframkallende, bl.a. hos planter. Plantepatogene bakterier trenger seg inn i plantevev gjennom naturlige åpninger eller skadd vev, og lever i første omgang av stoffer som lekker ut av plantecellene. Bakteriene kan påvirke dette med å danne toksiner og enzymer som gjør at plantecellene og cellenes funksjon endrer seg og dermed gir synlige symptomer. For eksempel er bløtråte vanlige symptomer i poteter, rotvekster og grønnsaker. Visnesyke kan være resultatet av bakterievekst i plantenes ledningsvev. Plantepatogene bakterier danner ikke hvilesporer, men har stort sett evne til å overleve ved å senke sin fysiologiske aktivitet til et minimum. På denne måten kan de tåle mer ekstreme forhold. Uten tilknytting til vertsplante-materialet har de fleste plantepatogene bakterier liten sjanse til å overleve i et miljø med andre, svært aktive mikroorganismer.
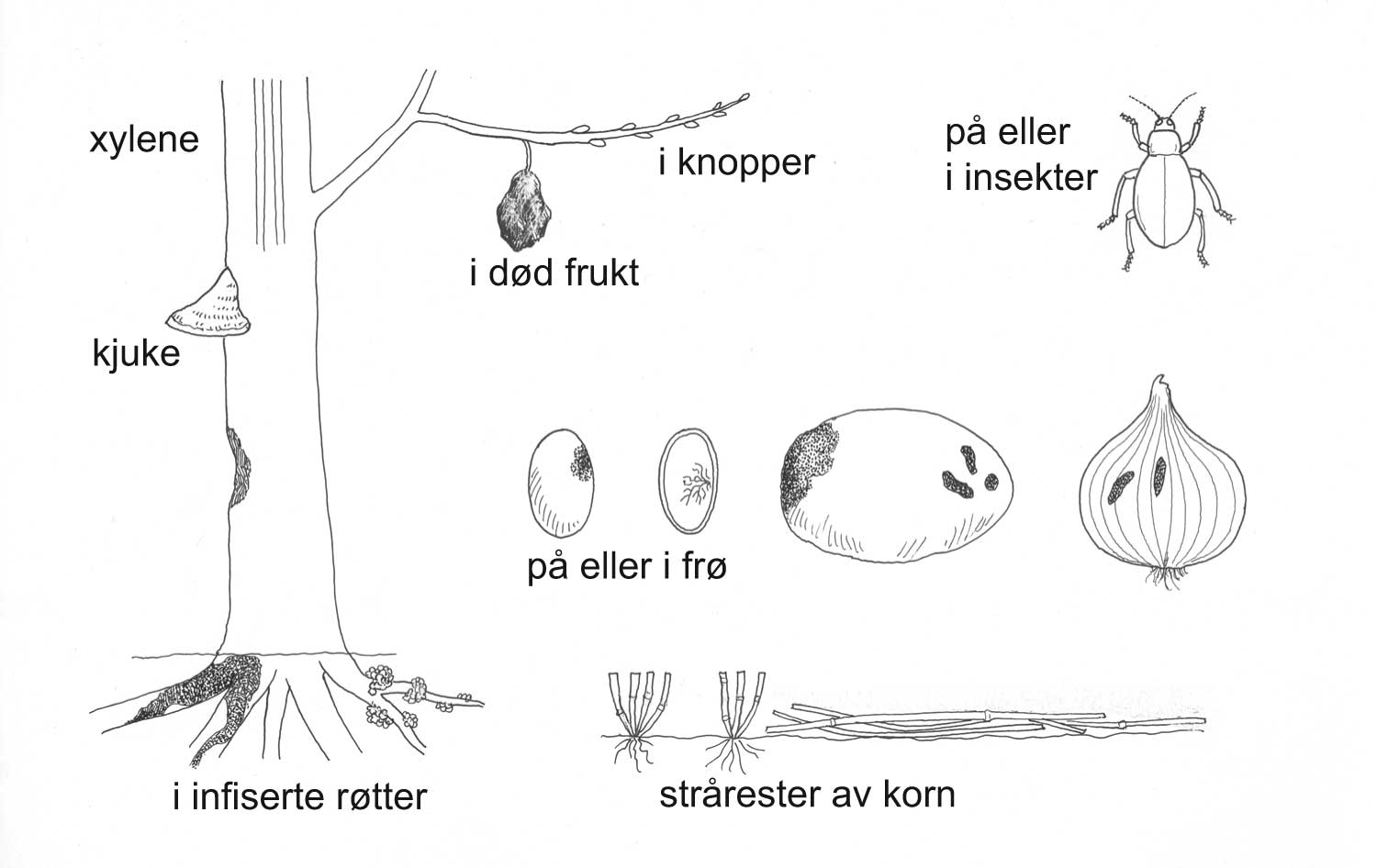
Virus er årsak til mange farlige sjukdommer, og alle virus er parasitter i celler, helt fra encellede organismer til høyerestående planter, dyr og mennesker. Virus skiller seg fra alle andre patogene organismer, ikke bare i størrelse og form, men også i sin enkle kjemiske og fysiske oppbygning, infeksjonsmåte, formering, hvor de opptrer i plantevevet og hva slags symptomer de fremkaller. De er oppbygd av en nukleinsyre (arvestoff, DNA eller RNA) med en proteinkappe omkring. Vi kan ikke si at virus er levende organismer på samme måte som bakterier og sopper. Virus har ikke noe eget stoffskifte, og kan ikke ta til seg næring eller formere seg på egen hånd slik bakterier og sopp gjør. Virus kan formeres i planteceller ved at de lurer cellene til å bruke sine enzymer til dette formål. Slik sett er virus egentlig en bit genetisk informasjon som bruker plantas biokjemiske apparat til å formeres opp for så å spre seg videre til nye celler og planter. Virus er derfor svært spesialiserte parasitter i plantene. Virus er så små at de ikke kan sees i vanlig lysmikroskop, men man må bruke et elektronmikroskop for å se og ta bilder av virus. Etter at et virus har brukt plantecellas apparat til å formere seg, kan de passere celleveggen og føres med væskestrømmen i ledningsvevet i plantene. Etter en tid vil alle deler av planta inneholde virus. Spredning fra plante til plante kan foregå med en vektor, som er fellesnavnet på levende organismer som overfører virus. Vektorer kan være insekter (for eksempel bladlus), sopp og nematoder. Virus kan også overføres ved at blader kommer i kontakt med hverandre eller skader seg (for eksempel at hår blir brutt av). Dette kan for eksempel skje ved påvirkning av vind eller ved kjøring i åkeren. Virus kan også overføres ved vegetativ formering av plantemateriale, for eksempel via settepoteter. I vegetativt formerte vekster er det derfor svært viktig med virusfritt utgangsmateriale. Frø er imidlertid mindre utsatt for virusinfeksjon, siden virus ikke så lett går over i frøet. Frøformerte kulturplanter er derfor vanligvis mindre utsatt for virus enn vegetativt formerte planter.
De tre gruppene mikroorganismer som til nå er omtalt, omfatter alle de viktigste sjukdomsfremkallende organismene i våre jord- og hagebruksvekster. Det finnes imidlertid to andre grupper med organismer som også kan fremkalle sjukdommer i planter. Disse kalles viroider og fytoplasma. Viroider er egentlig virus uten proteinkappe, dvs. kun biter med genetisk informasjon. Viroider har vært klassifisert som en egen gruppe mikroorganisme, men er nå klassifisert som virus. Fytoplasma er nærmest bakterier uten cellevegg. «Fyllodi» i blomster er et eksempel på en sjukdom fremkalt av fytoplasma. Blomsterfyllodi innebærer at det skjer en slags forgreining i selve blomsten slik at den får mange små og buskete utseende blomster der det normalt bare skulle vært en blomst.
Dersom vi betrakter verden fra sjukdomsorganismenes ståsted, vil deres liv være styrt av det samme instinktive «ønske» om formering og spredning som hos alle andre levende organismer. I dette ligger selvsagt svært ulike krav til omgivelsene, men innebærer for alle et viktig krav, nemlig tilgang på næring. Et plantepatogen finner maten pent servert i store mengder når vi i landbruket dyrker like vekster og arter over store områder og år etter år. Tilgang på næring er likevel ikke den eneste utløsende faktor for spredning og formering. Klimaforhold som temperatur, lys og fuktighet påvirker i høy grad utviklingen av en sjukdom. Plantenes resistens eller motstandskraft spiller også en stor rolle. Dette skal vi snakke mer om, men først skal vi se på hvordan noen av de sjukdomsfremkallende soppenes livssykluser kan arte seg.
Utbredelsen av en soppsjukdom på en plante eller i en åker, skjer vanligvis ved gjentagelse av følgende prosess: spiring - infeksjon - vekst - danning av en ny generasjon sporer. Dette kalles en infeksjonssyklus. Tida det tar for en syklus å fullføres betyr mye for hastigheten av sjukdomsutbredelsen. Det samme gjør mengden sporer som dannes i en syklus og sannsynligheten for at de nydannete sporene greier å starte en ny infeksjonssyklus. Det siste henger mye sammen med de mikroklimatiske forholdene som er til stede. Noen patogener fullfører bare en eller deler av en syklus i løpet av et år. Sotsopper på korn er eksempler på sjukdommer med bare en syklus per sesong. Etter at et korn med sotsmitte har spirt, infiserer sotsoppen planta etter hvert som den vokser. Infeksjonen kommer først til syne i akset etter skyting og fram mot modning. Ved riktige forhold spres sotsporene fra akset og over i nye friske aks, men soppen kan ikke danne nye sporer på disse nyinfiserte aksene. Infeksjonen kan ikke ses med det blotte øyet, og først neste sesong, når prosessen har gjentatt seg, kan man se resultatet av sporespredningen året før. For eksempel kan stinksot pga. stor sporeproduksjon oppformere seg kraftig fra en sesong til en annen dersom forholdene ligger til rette for det. Svært mange sopper har imidlertid flere (fra 2-30) generasjoner per år. Slike sjukdommer infiserer først plantene fra en primær smittekilde, for eksempel tørråte i settepotetknoller, mjøldogg som kommer med vind fra en infisert åker, eller grå øyeflekk fra infiserte planterester (figur 2.37). Under gunstige forhold dannes nye sporer på de nylig angrepne plantene, og slik kan sjukdommen spre seg videre fra stadig nye planter. Tenker man seg det enorme antall sporer som kan dannes på bare ett infeksjonssted, er det lett å forstå hvordan en sjukdom med mange sykluser kan bre seg eksplosivt i løpet av kort tid under de rette forholdene. Disse sjukdommene er med andre ord epidemiske.
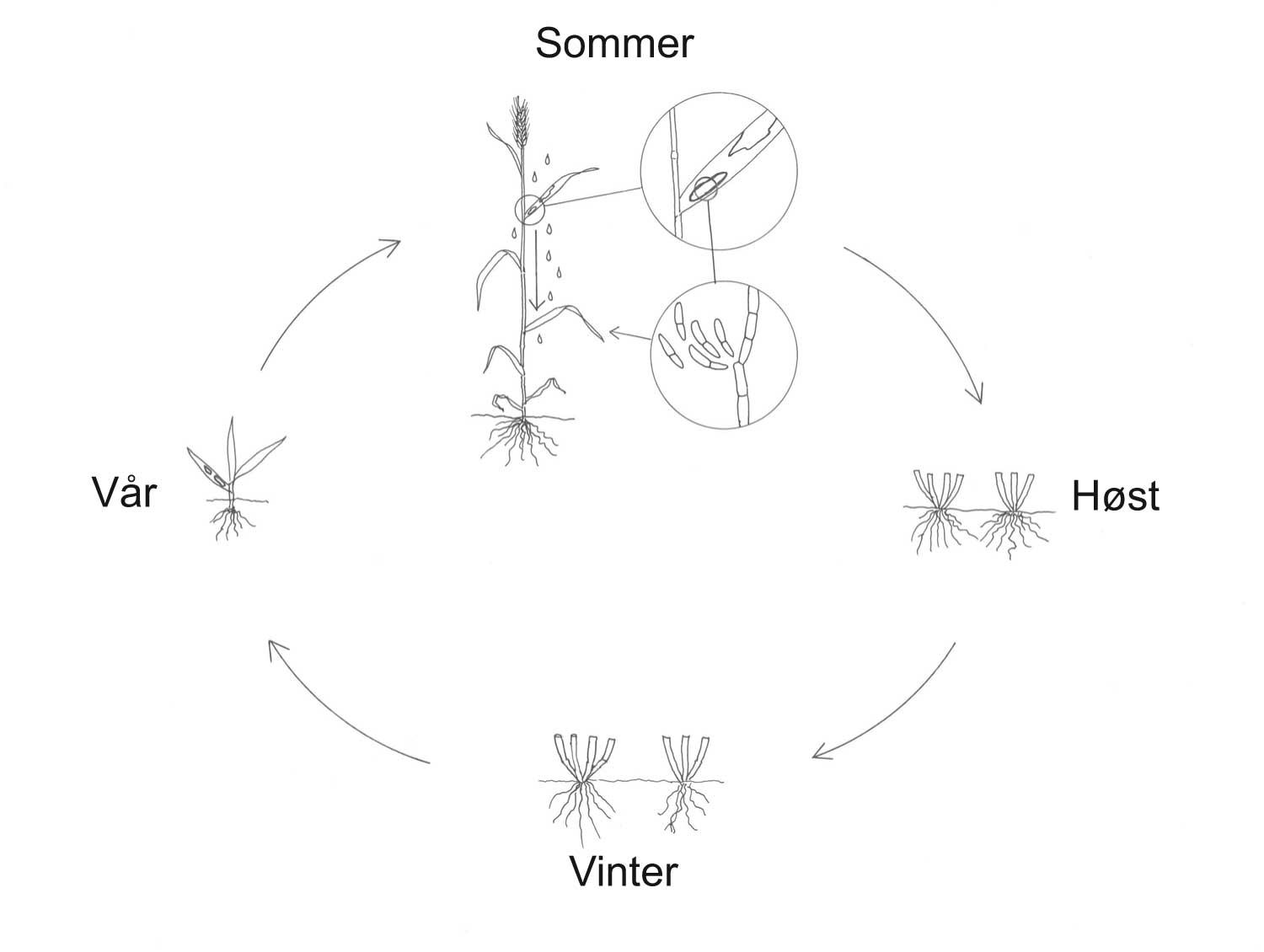
I prinsippet vil de samme forholdene være avgjørende for omfanget av en epidemi og omfanget av et angrep generelt. Det vil si at faktorer vi tidligere har nevnt som viktige for å få et angrep, vil også være viktige for utviklingen av en epidemi, men de riktige forholdene må i tillegg vare lenge.
Informasjon om disse faktorene er viktige for å forstå hvorfor et sjukdomsangrep utvikler seg som det gjør. Selv under tilsynelatende like forhold kan det være forskjeller i omfanget av et sjukdomsangrep, og kjenner vi de fleste faktorene kan vi kanskje lete oss fram til hvorfor det noen steder blir mindre sjukdom. Noe kan nok oppfattes som opplagt for de fleste, siden flere faktorer nærmest er allment kjent for de som dyrker jorda med interesse. Betydningen av vekstskifte eller valg av motstandsdyktige planter og betydning av fuktig vær er nok gammel kunnskap, men ikke mindre viktig å huske på.
Vi kan blant annet dele inn faktorene på følgende måte:
Vertsplantefaktorer
Patogenfaktorer
Miljøfaktorer
Dyrkingspraksis/menneskelig påvirkning
Hvordan planter kan unngå sjukdom
Noen planter blir lettere angrepet av sjukdom enn andre. Evna til å motstå eller overvinne angrep av et patogen, kalles resistens. På samme måte som ulike sorter varierer i resistens overfor for eksempel en patogen sopp, kan soppen variere i aggressivitet overfor planten. Dette kalles virulens. Selv planter som i utgangspunktet er mottakelig for et bestemt patogen, vil ha en viss grad av noe vi kaller for uspesifikk resistens mot «sine» patogener. Dette kalles ofte for generell resistens, eller feltresistens. Som navnene tilsier, beskytter ikke denne resistensen planta fra å bli infisert, men bremser utviklingen av sjukdommen på de enkelte plantene. På den andre siden har vi spesifikk resistens, som kan gjøre en plante helt eller svært motstandsdyktig mot angrep av et bestemt patogen. Spesifikk resistens bestemmes av få gener mens uspesifikk resistens bestemmes av mange gener. Dette gjør at spesifikk resistens lettere vil kunne overkommes av patogenet, og planta vil kunne miste sin resistens. Uspesifikk resistens, eller feltresistens er ikke fullstendig, men til gjengjeld mer robust og varig.
En plante som i utgangspunktet er mottakelig, kan av og til «unnslippe» en sjukdom. Dette kan skje på grunn av den enkelte plantes fysiologiske egenskaper. Frøene kan for eksempel modne tidligere enn plantene rundt, eller herdingen skje tidligere enn andre planter av samme sort. Alder spiller ofte en rolle siden noen planter bare er mottakelige for et patogen i visse stadier av sin utvikling. I mange tilfeller unnslipper plantene sjukdom fordi de vokser i blanding (sortsblandinger, artsblandinger) med andre ikke-mottakelige planter. Mengda inokulum eller smittestoff som når fram til disse plantene blir mindre enn hvis de vokser i monokultur.
Mange planter unnslipper et angrep av Pythium og Phytophthora dersom temperaturen er høy og jordfuktigheten lav, mens for eksempel hvete unngår liknende sjukdommer forårsaket av Fusarium og Rhizoctonia hvis temperaturen er lav. Den vanligste grunnen til at sjukdom ikke utvikler seg er manglende fuktighet i form av regn eller dugg. I såkalte tørre år er utbrudd av tørråte, epleskurv og andre sjukdommer langt færre enn i fuktige år. Patogenene trenger ofte en vannfilm på planten, eller høy luftfuktighet, i nesten alle stadier av sin livssyklus. Vi har imidlertid også eksempler på patogener som utvikler seg bedre under tørre forhold, og som hemmes av regn eller vanning. Streptomyces scabies, eller flatskurv på potet, er eksempel på en slik sjukdom som man altså kan «vanne » seg bort fra. Dette er fordi bakteriell antagonisme mot flatskurv fungerer for dårlig under tørre forhold. En annen faktor er pH. Klumprotsoppen blir hemmet av høy pH, mens nevnte flatskurvsopp hemmes av lav pH.
Enkelte sortsegenskaper hos plantene kan gjøre at de unnslipper sjukdom selv om de for øvrig er mottakelige. For eksempel slipper tidlige potetsorter som regel lettere unna tørråte enn sene sorter. Dette er fordi knolldanningen allerede er i gang før tørråteinfeksjonene setter inn. Tidlige sorter er faktisk ofte mindre resistente enn sene sorter, men unngår altså lettere infeksjon pga. sin tidlighet. Rask vekst kan også gjøre at plantene «vokser fra» et sjukdomsangrep. Hvis vi bruker disse forskjellene bevisst, kan det være med på å begrense sjukdomsomfanget hos den enkelte dyrker, i den enkelte åker osv.
Symptomer på sjukdomsangrep kan for eksempel være tydelige flekker, spesifikke for en sjukdomsorganisme, noe som gjør det enkelt å stille diagnose. Andre kan være diffuse og lite særpregede. Ved usikkerhet kan det være nødvendig å ta i bruk mikroskop eller annet teknisk utstyr eller metoder for sikkert å kunne fastslå hva som er årsaken. Det finnes mye godt bildemateriale og diagnoseverktøy for de som vil lære å kjenne sjukdomssymptomer fra hverandre. Her i boka skal vi omtale symptomer mer generelt.
Det finnes typer symptomer som ofte skyldes en gruppe sjukdomsorganismer. For eksempel vil en svulst eller bløtråte som regel være forårsaket av en bakterie, mens rust og mjøldoggbelegg er dannet av sopper. Tabell 2.9 viser hvilke symptomer de ulike sjukdomsorganismene kan lage, mens figur 2.38 viser hva som menes med noen av symptomnavnene som nevnes i tabellen.
Tabell 2.9 Symptomer på sopp, virus og bakterier
| Sopp | Virus | Bakterier | |
| Bladflekker | X | (X) | X |
| Avdøde plantedeler (nekroser) | X | X | X |
| Sotsporeklumper | X | ||
| Rustflekker | X | ||
| Svulster | X | ||
| Visning | X | X | |
| Redusert vekst | X | ||
| Vekstforstyrrelser | X | X | |
| Fargeendringer/kloroser | X | X | (X) |
| Tørrråte/tørre råter | X | ||
| Bløtråte/bløte råter | X | X |
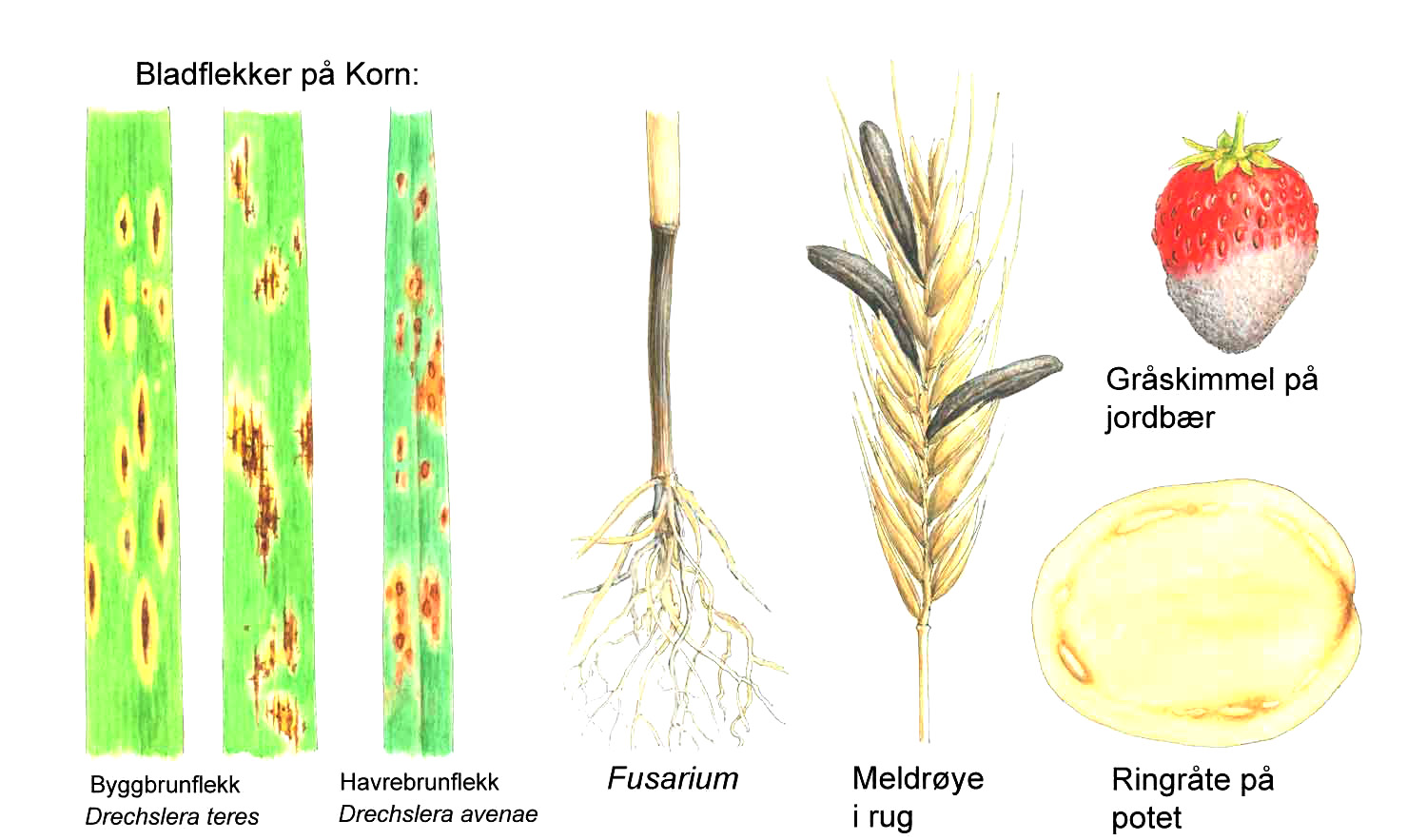
Enkelte av de nevnte symptomene kan også skyldes andre skadegjørere, for eksempel kan redusert vekst skyldes nematodeangrep.
Sopp, virus og bakterier gir mer eller mindre ulike symptomer på plantene. Angrep av en skadegjører er imidlertid ikke det eneste som kan gi symptomer. Næringsmangel kan i en del tilfeller gi liknende symptomer som for eksempel et soppangrep. Å kunne skille symptomer på sjukdom fra andre typer stress og mangler er avgjørende ved valg av videre strategi.
Noen eksempler: Et angrep av en levende organisme på en grønn plantedel, som et blad, vil som regel føre til en bleking av bladet, eller «klorose» som dette kalles. I neste omgang vil den angrepne delen kunne dø og bli brun. Dette kalles nekrose. Næringsmangel vil også kunne gi nekrotiske flekker på bladene, men disse vil ofte opptre i et mer regelmessig mønster enn tilfellet er for en levende skadegjører. Det er imidlertid vanskelig å gi generelle regler fordi det alltid finnes unntak.
Å kjenne de ulike symptomene fra hverandre kan selvsagt virke som en omfattende og vanskelig oppgave. Med noe kunnskap om grunnleggende og viktige symptomer og kjennetegn har man imidlertid en god basis.
Økologisk produksjon i seg selv er ikke en garanti mot sjukdommer, men forebyggende tiltak og strategier preger denne driftsmåten sterkere enn det som er vanlig i konvensjonell produksjon.
Økologiske gårdsbruk vil som regel ha en mer allsidig drift enn konvensjonelle gårdsbruk. Typiske vekstfølgesjukdommer (sjukdommer som kommer av dårlig vekstskifte) vil være mindre problematiske ved allsidig drift og godt vekstskifte. Sjukdommer som følger såfrø og såkorn vil derimot kunne bli mer problematiske ettersom muligheter for direkte behandlingsmetoder i form av beising er mer begrenset. Pløying er nok mer utbredt ved økologisk drift enn ved konvensjonell drift i dag. Pløying begraver planterester og kan begrense spredning av enkelte sjukdommer som overlever i slike rester. Slik kan man fortsette å se på enkelt-tiltak og i hvilken grad disse kjennetegner driftsformer og samtidig påvirker sjukdomssituasjonen.
Allsidigheten varierer i dag mye både innenfor det økologiske og innenfor det konvensjonelle landbruket. Forskjeller i sjukdomssituasjonen vil eksistere mellom de fleste gårdsbruk som følge av alle ulike forhold som er med å påvirke sjukdomsutviklingen. Selv om man kan si noe svært generelt om sjukdomssituasjoner på økologiske og konvensjonelle gårder, er det til syvende og sist driftsopplegget sammen med gårdens naturgitte forhold som avgjør. En allsidig drift med gode omløp vil i stor grad ha betydning for sjukdomssituasjonen enten det gjelder økologiske eller konvensjonelle bruk. Slikt sett er det kanskje viktigere å se på hvordan sjukdommer blir påvirket av konkrete dyrkingstekniske tiltak, for eksempel omløp/vekstskifte, jordart, næring (type og mengde) og jordarbeiding. Dette blir nærmere omtalt i neste kapittel av boka.
«Hvilken vidtløftig og vanskelig oppgave det ofte kan være at få undersøgt rigtig grundigt de forskjellige smådyrs hele livs- og udviklingshistorie, der ved sin levevis får økonomisk betydning som skadedyr for vore kultur planter.» Konservator W. M. Schøyen 1888
For å forstå insekters og andre dyrs opptreden som skadedyr er det nødvendig med en enkel innføring i hvordan skadedyrene er bygd opp, hvordan de opptrer, overvintrer, oppformerer seg, finner vertsplantene og gjør skade.
Når man omtaler skadedyr på planter i økologisk landbruk er det først og fremst insekter det tenkes på. I tillegg kommer nematoder (se eget avsnitt lengre bak), midd, snegler, smågnagere og storvilt. Pattedyrene skal ikke behandles i denne boken. De insektsordnene som er mest aktuelle som skadedyr er (rangert fra mest primitive til mest utviklete) saksedyr, trips, plantesugere, biller, sommerfugler, tovinger (fluer) og veps (planteveps). Vi omtaler ofte midd sammen med insekter. Midd er edderkoppdyr, men i likhet med insekter tilhører midd overordenen Arthropoda, leddyr.
Skadedyrene kan grupperes etter type munndeler og type skade. De kan gjøre skade enten under egglegging, spising eller fordi de påfører planten sjukdomsframkallende organismer som bakterier, virus, o.l. I tillegg er det noen få arter som krøller bladene rundt seg for ly, og dette kan skade plantene, spesielt hvis vekstpunktet blir dekket over. Noen arter stikker eggleggingsrøret inn i plantevevet, og dette kan gi små prikker på plantene og eventuelt være inngangsporter for plantesjukdommer. Munndelene har ulik utforming hos ulike skadedyr. Stikkende-sugende munndeler finner vi bl.a. hos bladlus og teger. Disse suger plantesaft. Skaden kan være bladkrusing og dødt plantevev. Hos noen arter blir et toksin som dreper plantecellene spyttet inn i planten, og dette kan føre til at bl.a. vekstpunktet dør eller deler seg på en unormal måte. Flere av disse sugende skadedyrene utskiller en sukkerholdig væske (honningdugg) som gir grobunn for svertesopp på planteflaten, noe som går ut over både fotosyntese og produktkvalitet.
Både trips, midd, snegler og nematoder spiser på en måte som kan kakteriseres som suging av plantesaft. Fra trips, midd og snegler er skaden oftest celledød som er synlig på planteflatene. Noen ganger kan snegleskade gå dypere i plantedelene slik at det ligner gnagskade. Nematodeskade vil oftest oppfattes som svak vekst eller visning, og det er vanlig å overse et angrep før nematoder er funnet i prøver.
Minering i plantene, som sees som ganger eller «tunneler» i plantedelene, gir forskjellige typer skade avhengig av hvilken plantedel det er i. Det er oftest larver av sommerfugler, fluer og noen biller som minerer i planter. Hvis det er i røttene kan det føre til visning og evt. plantedød. I andre plantedeler kan det forårsake forringet kvalitet, råte, og hvis angrepet er sterkt, kan planten dø eller avlingen utebli.
Den skaden som er mest iøynefallende er gnagskade. Insekter med bitende-gnagende munndeler gnager tvers gjennom plantematerialet. Som regel gir dette redusert bladareal for fotosyntese eller direkte skade på produktet. Noen ganger kan plantene snauspises eller ødelegges totalt fordi de blir gnagd av ved jordoverflaten. Insekter som gnager er vanligvis larver av sommerfugler, planteveps eller biller, eller voksne biller (tabell 2.10).
Tabell 2.10 Munndelenes utforming hos noen skadedyr og skaden de forårsaker (se også figur 2.39)
| Gruppe | Larve | Voksen | Skade |
| Saksedyr | Bitende-gnagende | Bitende-gnagende | Flisete |
| Teger | Stikkende-sugende | Stikkende-sugende | Celledød og veksthemming (toksin i spyttet) |
| Bladlus | Stikkende-sugende | Stikkende-sugende | Bladkrølling og virussmitte |
| Kvitfly | Stikkende-sugende | Stikkende-sugende | Suger ut plantesaft og svekker bladene |
| Sikader | Stikkende-sugende | Stikkende-sugende | Suger ut plantesaft og svekker bladene |
| Skjoldlus | Stikkende-sugende | Stikkende-sugende | Suger ut plantesaft og svekker plantene |
| Sugere | Stikkende-sugende | Stikkende-sugende | Bladkrølling |
| Trips | Skrapende-sugende | Skrapende-sugende | Flekker og striper |
| Sommerfugler | Bitende-gnagende | Sugesnabel (ikke skadedyr) |
Miner og hullgnag |
| Tovinger | Bitende-gnagende munnhakker | Stor variasjon, alle lever av flytende føde (ikke skadedyr) | Miner og ganger, celledød og veksthemming fra gallmygglarver |
| Bladveps | Bitende-gnagende | Ofte bitende-gnagende | Hullgnag |
| Biller | Bitende-gnagende | Bitende-gnagende | Hullgnag, noen minerende larver |
| Midd | Stikkende-sugende | Stikkende-sugende | Avfargete flekker |
| Nematoder | Munnbrodd | Munnbrodd | Celledød og forvridd vekst |
| Snegler | Raspetunge | Raspetunge | Skråkantete og flisete hull |
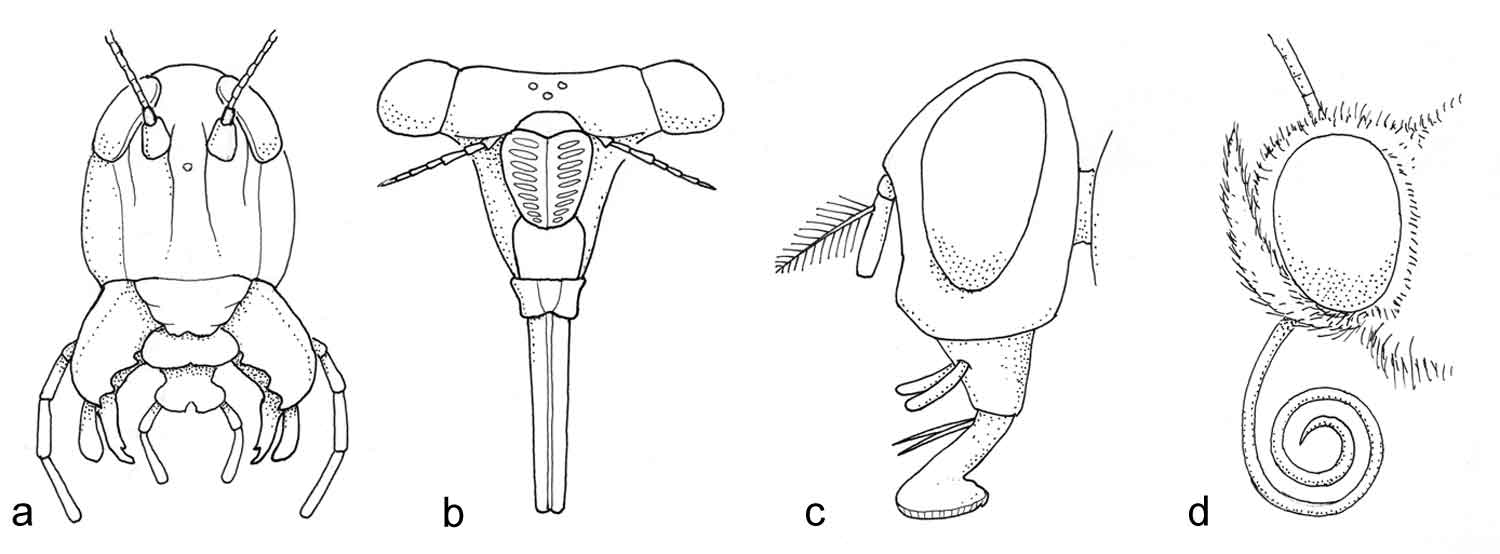
Nematoder
Nematoder (rundormer) er virvelløse dyr der kroppsvæsken fungerer som skjelett (hydrostatisk skjelett). Det osmotiske trykket gir kroppen stødighet og form, og muliggjør nematodenes svømmende bevegelsesmåte. De har de samme organsystem som finnes hos andre dyr, men mangler hjerte- og pusteorgan. Nematoder finnes i alle biotoper over hele jordkloden og er den største gruppen av flercellede dyr. De fordeler seg over 250 familier som igjen grupperes i nærmere 20 ordener, hvor orden Tylencida omfatter det største antallet nematoder som lever på planter. En næringsøkologisk basert inndeling for disse nematodene er:
Mer utdypende informasjon om nematoder som skadedyr finner du i tekstboks 2.16.
Av kjente nematodeslekter er ca. 7 % parasitter på planter og 41 % parasitter på dyr, mens resten er frittlevende med forskjellige levesett. Om lag 2200 nematodearter lever på planter, men under nordiske forhold er kanskje bare 100 av disse skadelige i ordets rette forstand. Det store flertallet kan snarere benevnes som vekstspisere, som tar seg en «jafs» av plantene uten at veksten lider nevneverdig av det.
Nematoder finnes over alt, og i en kvadratmeter jord kan det finnes millioner av dem. Vanligvis er de mikroskopiske, trådformede dyr som lever av døde dyre- og planterester i jord eller sediment, eller som parasitter på dyr, mennesker og planter. Omfanget av skader forårsaket av nematoder som lever på eller inne i planter er avhengig av nematodeart, antall individer, planteslag, jord- og klimaforhold. Angrep reduserer avling og kvalitet, og kan forkorte vekstens levetid. Plantevev som er skadet av nematoder er mer utsatt for sjukdom forårsaket av sopp og bakterier. Dessuten kan enkelte nematoder overføre virus til planter. I felt er det vanlig å finne flere arter av skadelige nematoder i det samme område. I jordbruksområder, hvor dyrkingen må anses som spesialisert, finner en ikke samme balanse mellom nematoder og planter som det man gjør i uberørt natur der nematoder gjennom mer enn 400 millioner år har utviklet seg sammen med sine respektive vertsplanter.
De fleste nematoder som finnes i jord- og plantematerialer er 0,3-1,0 mm lange, men noen kan også bli så lange som 5-7 mm. I hovedsak er de gjennomsiktige og dermed vanskelig å se uten bruk av mikroskop. Unntaket er cystenematoder hvor hunnene fra om lag midten av vekstsesongen kan sees med det blotte øye på rotsystemet hos en del planteslag. Velkjent er potetcystenematoder (potetål) som kan sees på potetrota som knapt millimeterstore runde hvite og gule kuler fra cirka 40 dager etter setting. For cystenematoder på korn tar det normalt 70 dager fra såing før man ser de hvite små kulene på rotsystemet.
Særlig hos arter av cystenematoder og stengelnematoder forekommer flere raser, også kalt patotyper. Nematoder av ulike raser innen en art er utseendemessig helt like. For å skille de forskjellige rasene benyttes derfor spesielt utvalgte testsorter som er tilpasset nematodearten. Rasebestemmelse gir kunnskap om sorter av et planteslag er mottakelig for bestemte populasjoner av nematoder, eller om sorten er motstandsdyktig (resistent). Dette er viktig kunnskap for dyrkeren med tanke på riktig sortsvalg for å oppnå så god avling som mulig.
Generelt gjennomgår nematoder fire ungstadier og hudskifter fra egg til de blir voksne. Det er andre ungstadiet som klekker fra egget. For å trenge gjennom celleveggene, enten for å krype inn i planta eller bare for å spise, har nematoden en spiss munnbrodd. Denne brodden er et spesielt kjennetegn for gruppen som tilhører nematoder som spiser på levende planter. Hver art av nematoder har sin spesielle livssyklus. Under våre klimatiske forhold er det noen nematoder som i løpet av sommermånedene rekker å ha flere generasjoner, for eksempel rotsårnematoder. Men det finnes også de som bare rekker én generasjon i året, slik som cystenematoder. De fleste nematoder legger sine egg direkte i jorda. Noen tar imidlertid bedre vare på eggene sine for å øke sjansen for arten å overleve. Flere nematoder har ekstrem overlevelsesstrategi. Benevnelse som kryptobiose (hemmelig liv), anabiose («gjenoppstandelse»), eller anhydrobiose, er kjente begreper. Som det siste navnet antyder, tilpasser nematodene seg tørke gjennom selv å tørke ut slik at all fysiologisk aktivitet stopper, for så å «vekkes til liv» når ytre faktorer, først og fremst vannrelaterte, blir ideelle. I denne tilstanden kan nematodene overleve i svært lang tid, faktisk flere titalls år, og utstå ekstreme påvirkninger. Potetcystenematoder er en slik art. Her legges eggene inne i hunnens kropp, og når hun dør, omdannes skinnet til et ganske motstandsdyktig skall. Selv uten vertsplante i mer enn 40 år har man fått klekking fra slike cyster.
Nematoder er ikke bare skadelige. Noen kan anvendes som modellorganismer i utviklingsbiologisk forskning, som bioindikatorer på miljøforandringer, og ved biologisk bekjempelse av insekter og snegler. (se Kapittel 4 om biologisk kontroll)
Nematoder er i hovedsak enkjønnet. Flere planteparasittære nematoder, for eksempel rotgallnematoder, har kjønnsløs forplantning i form av jomfrufødsel.
Symptom på skade av nematoder viser seg ofte som flekker med misvekst i åkeren. På plantene er ikke alltid symptomene like entydige, og krever ofte hjelp av mikroskop.
Siden nematodene i seg selv er stasjonære, må de ha hjelp til forflytting over lengre avstander. Hjelpen kan komme fra vind, flomvann, jordarbeid, redskap, transport, plantesalg, m.m., men som hovedregel er det i forbindelse med at jord flyttes på et eller annet vis.
Bekjempelse av nematoder er i mange tilfeller vanskelig. Naturlige fiender som spesielle sopper, bakterier, rovnematoder, o.a. kan finnes i jorda, men det gjenstår ennå å få slike system til å fungere maksimalt med tanke på å hindre avlingstap. Blant annet solarisering, varmebehandling, damping, drukning, brakking, og valg av ikke-vertsplanter kan være tiltak for å redusere nematodebestand. Frost kan også gjøre at en del nematoder fryser i hjel, men for eksempel for potetcystenematoder må man ned under -30 ºC før det blir noe hjelp av betydning. Forebyggende tiltak som renhold, valg av riktig plantemateriale, vekstskifte, og analyse av jordprøver for nematoder er svært viktig for å redusere og hindre skade av nematoder i plantekulturen.
Tabell 2.11 Viktige planteskadelige nematoder
| Norsk navn | Noen planter som skades |
| Potetcystenematoder | Potet |
| Korncystenematoder | Korn, gras |
| Kløvercystenematoder | Kløver, belgvekster |
| Grascystenematoder | Gras |
| Kroknematode | Korn, gras |
| Stengelnematode | Frø, løk, purre, kløver, blomsterløk |
| Potetråtenematode | Potet, narcisser, gulrot |
| Furuvednematode | Bartrær |
| Rotgallnematoder | Potet, grønnsaker, hagebruksvekster |
| Rotsårnematode | Potet, grønnsaker, frukt, bær, korn, gras |
| Bladnematoder | Jordbær, veksthusplanter |
| Stuntnematoder | Korn, gras |
| Torvnematode | Roser, veksthusplanter |
| Nålnematoder1) | Jordbær, gras, grønnsaker |
| Dolknematoder1) | Frukt, bær, veksthusplanter |
| Stubbrotnematoder1) | Potet, grønnsaker |
1) Overfører også virus.
Insektfysiologi
Hvis vi skal bekjempe insekter som er skadedyr kan det være en fordel å kjenne til deres oppbygging slik at vi kan utnytte de svakhetene som finnes. Ytterst har insekter et tynt lag som består av vokslignende stoffer som er syrebestandige. Dette gir beskyttelse mot inntørking, fuktighet og noen sjukdomsorganismer. Under dette laget er det to porøse lag som inneholder kitin som gir beskyttelse mot vanlige etsende kjemikalier. Under kitinlaget er selve huden og under den er en membran. Til sammen danner alle disse lagene det som tilsvarer skjelettet hos insektet.
Insektskroppen er delt opp i flere segmenter (ledd), og mellom disse mangler det harde, ytre laget som ellers dekker leddene. Dette gjør kroppen bevegelig. De myke partiene kan også være svake punkter der f.eks. sopp-patogener kan få fotfeste. I tillegg er det mange åpninger gjennom huden i forbindelse med kjertler og pustehull og disse kan brukes av bl.a. nematoder som angriper insekter (se figur 2.40).
Sanseorganer som brukes i forbindelse med utvelgelse av føde (planter) er lukt-, syn- og smaksorganer. Det er oftest voksne hunner som søker seg fram til plantene som skal være mat til avkommet. Lukteorganene sitter hovedsakelig i antennene hos voksne insekter (figur 2.41) og i munnområdet hos larver.

Synsorganene er alltid på hodet og består av punktøyne og fasettøyne (figur 2.42a). Punktøynene oppfatter lys og mørke, mens fasettøynene bl.a. oppfatter bevegelse, avstand og farger. Fasettøynene er mer utviklet hos predatorer som skal fange byttet enn hos jordboende plantespisere som f.eks. rotlus.
Smaksorganene brukes til å bestemme om maten (f.eks. en plante) er riktig eller ikke. De fleste smaksorganene sitter i og rundt munndelene, men hos flere arter sitter de også på føttene (figur 2.42b). Dette gir insektet muligheten til å smake på maten raskt etter landing og godta eller avvise den uten å bruke mye tid. I tillegg har noen arter smaksorganer i eggleggingsrøret som brukes til å velge hvor eggene skal legges.
Insekter som lever på planter har utviklet seg slik at de får feste på plantene. Mange arter har spesielle strukturer på føttene som gir dem feste (figur 2.43). Dette kan være kroker, hår, sugekopper eller kjertler som produserer klebrige stoffer som gir en «post-it»-effekt. Det er dette som gjør at dyrene kan gå opp en rett vegg eller stå opp ned i taket.
Økosystemet på dyrket mark
Å forstå hvordan et enkelt individ opptrer på laboratoriet er ikke nok til å forklare atferden på dyrket mark. Dyrene lever sammen i populasjoner og det er egenskapene til populasjonene som betyr mest for skadeomfanget. Skadedyrenes populasjoner er avhengige av andre dyr, vertsplantepopulasjoner, klima, jordsmonnet og terrenget. Målet med skadedyrstudier er å forstå hvordan økosystemet som skadedyrpopulasjonen er en del av, fungerer. Denne forståelsen vil hjelpe oss til å påvirke utviklingen av skadedyrpopulasjonene slik at vi unngår økonomisk skade ved å gripe inn i økosystemet.
Økosystemet på dyrket mark utsettes for mange forstyrrelser. Pløying, harving, o.l. «stiller klokka» tilbake hvert år. Dette påvirker faunaen sterkt i forhold til mer uforstyrrede naturlige økosystemer. Ikke minst ettårige kulturer, som for eksempel korn, er ofte dyrket på en måte der jorda ligger brakk etter høsting. Dette reduserer både mengda og mangfoldet av dyreliv på det stedet. I en klassisk undersøkelse i England ble det for eksempel funnet ti ganger så mye leddyr i naturlig grasmark som i en kornåker. Også slåtten reduserer mengden leddyr og reduksjonen er større hos nyttedyr enn skadedyr (figur 2.44).
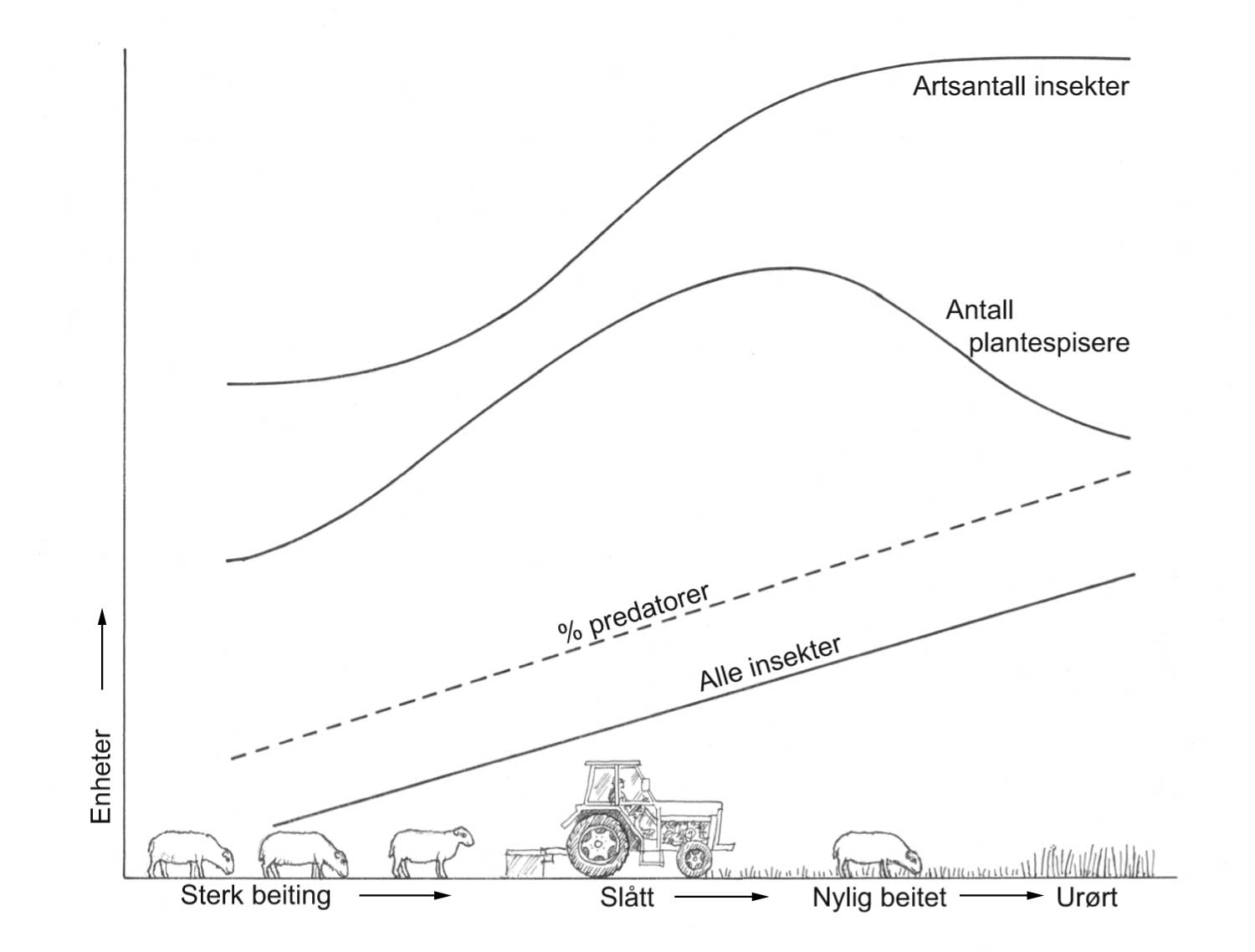
Den korte tida vi dyrker åkerkulturer fører også til lavere artstall av dyr. Dette gjelder mest for rovdyr og parasitter. Antall plantespisende arter blir også redusert, men antall individer pleier å øke sterkt. Dette gjør det vanskelig å bygge opp en balanse mellom skadedyr og nyttedyr i kortvarige kulturer.
Plantene som vi finner i et naturlig økosystem er et produkt av naturlig seleksjon. I en plantekultur er det derimot mennesker som velger plantene. I unge kulturer av dyrkede planter er det ofte mye åpen jord mellom plantene. Dette kan påvirke angrep av skadedyr fordi mange av disse reagerer på kontrast mellom plante og åpen jord når de leter etter vertsplanter. Langvarige samspill mellom dyr og planter i naturlige økosystemer fører til reduserte svingninger i populasjonene, og dette gir økende stabilitet. Når mennesker velger og dyrker planter er samspillet mellom dyr og plante forholdsvis nytt, noe som fører til mindre stabilitet og større sjanser for bølger av skadedyrangrep.
Ved å velge vegetasjonen påvirker dyrkeren økologien på mange måter, som i sin tur påvirker skadedyrsituasjonen. Det er ikke bare valg av plante, men sort, vekstfølge, størrelsen på åkeren, valg av nabovekster, m.m. som også spiller inn.
Som oftest er skadedyr som angriper kortvarige vekster svært mobile. Da kan de finne fram til kulturveksten tidlig, ofte før sine naturlige fiender. Kulturvekster består av ensartede plantebestander som har mistet mye av sitt forsvar. I naturlige økosystemer vil plantene i tillegg til egne forsvarsmekanismer ha forsvar pga. variasjonen i plantebestanden i utviklingsstadium, blanding med andre vekster og genotyper.
Det er viktig å være klar over at når mennesker dyrker planter, så skaper vi et nytt økosystem på stedet. De naturlige, regulerende kreftene som gjør at plantespisere sjelden herjer i naturen er ikke lenger til stede. I tillegg, jo mer kortvarig kulturen er, jo vanskeligere er det å få hjelp av de naturlige økologiske faktorer mot skadedyr. Derfor må vi dyrke på en måte som etterligner naturen så langt som mulig, mens vi samtidig må kunne utføre praktiske tiltak.
Overvintring
Insekter har forskjellige strategier for å overleve den norske vinteren. Det de har til felles er tilnærmet nullaktivitet. Ulike arter overvintrer i ulike stadier (egg, larve, puppe, voksen). Noen overvintrer på det plantematerialet der de utviklet seg. Andre finner beskyttelse under løv, skogsstrø eller under bakken (figur 2.45).
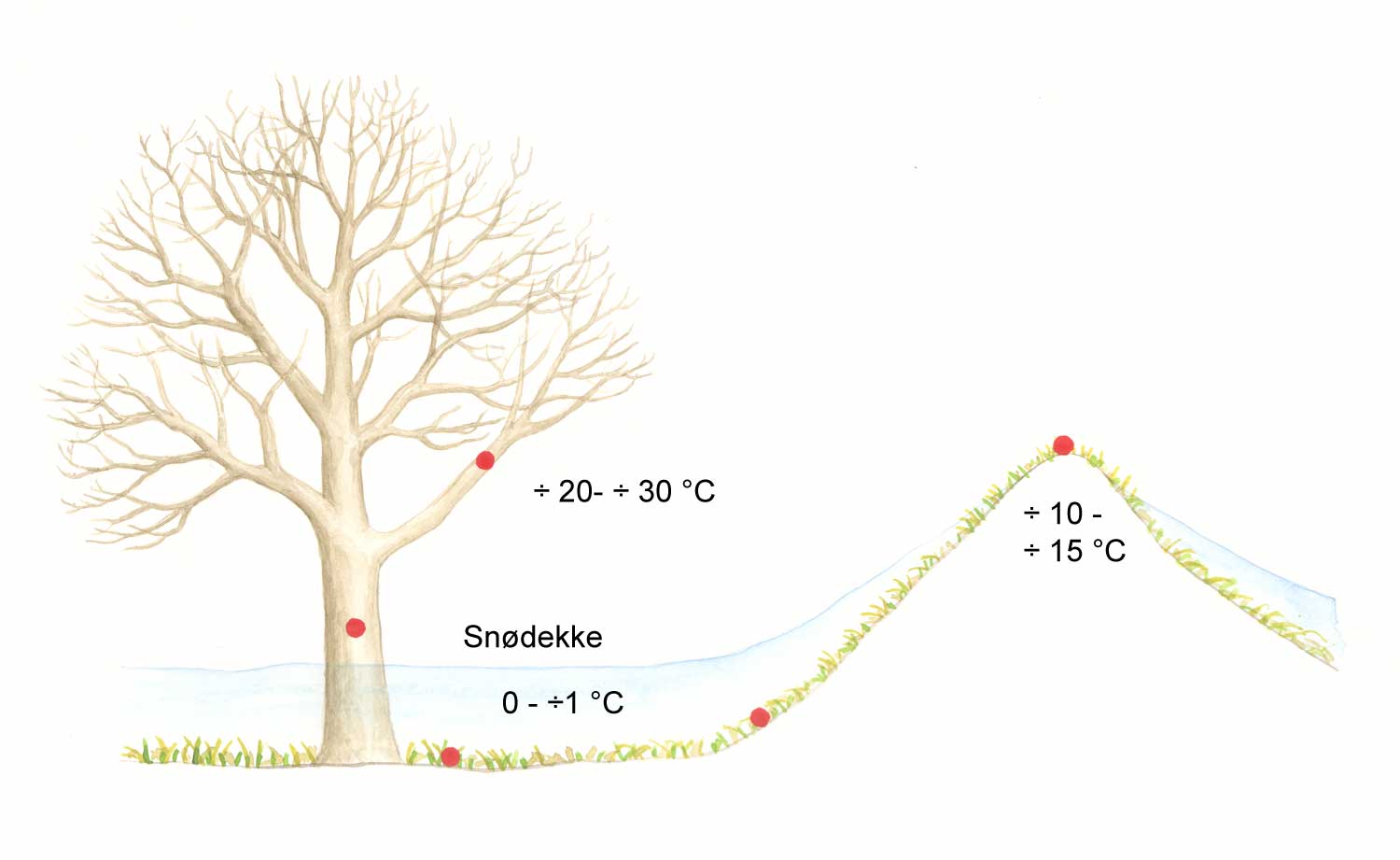
Insekter som ikke er tildekket mot kulda må beskytte seg mot frysing ved hjelp av forandring i innholdet i kroppsvæsken. En del insekter kan danne alkoholholdige stoffer som fungerer på samme måten som frostvæske i bilmotorer. Da tåler insektene temperaturer helt ned i -50 ºC. Insekter som har tvungent vertsskifte, som f.eks. noen bladlusarter, tilbringer vinteren på vertsplanten som de skal begynne den nye sesongen på. Disse kan bare sette i gang å spise der de er når temperaturen stiger nok om våren. De fleste andre må oppsøke vertsplanter for spising eller oppformering.
Et insekt blir aktivt (for eksempel klekker fra egget) om våren ved en bestemt temperatur, mens utvikling til neste stadium krever en bestemt varmesum. Dette gjelder også for videreutvikling og vekst. Hvis vi vet hva varmesumskravet er for en art kan vi beregne når arten vil opptre og angripe plantene. For enkelte skadedyr er det utført detaljerte biologiske studier som viser hvordan utvikling av de forskjellige stadiene er påvirket av temperaturen (figur 2.46).
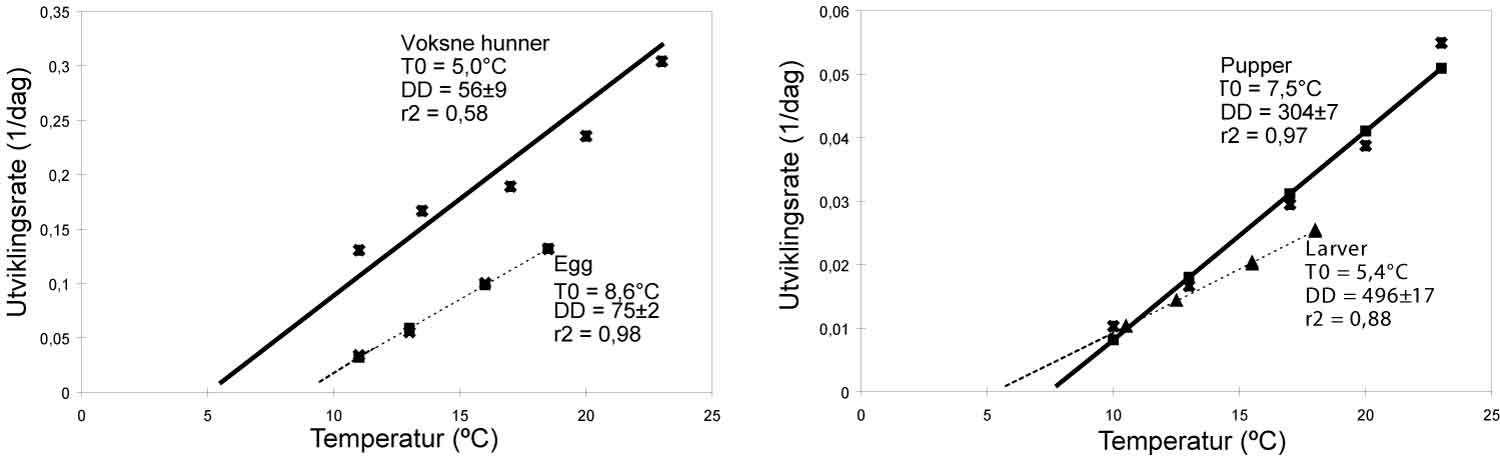
Oppsøking av vertsplanter
De insektartene som opptrer som skadedyr i Norge har oftest bare én eller noen få generasjoner hvert år. Når vi tar i betraktning at vekstskifte hører naturlig til økologisk landbruk, betyr dette at insektene må oppsøke vertsplantene fra det stedet hvor de overvintret. Dette gjelder både for insekter som overvintrer i og utenom kulturen (figur 2.47).
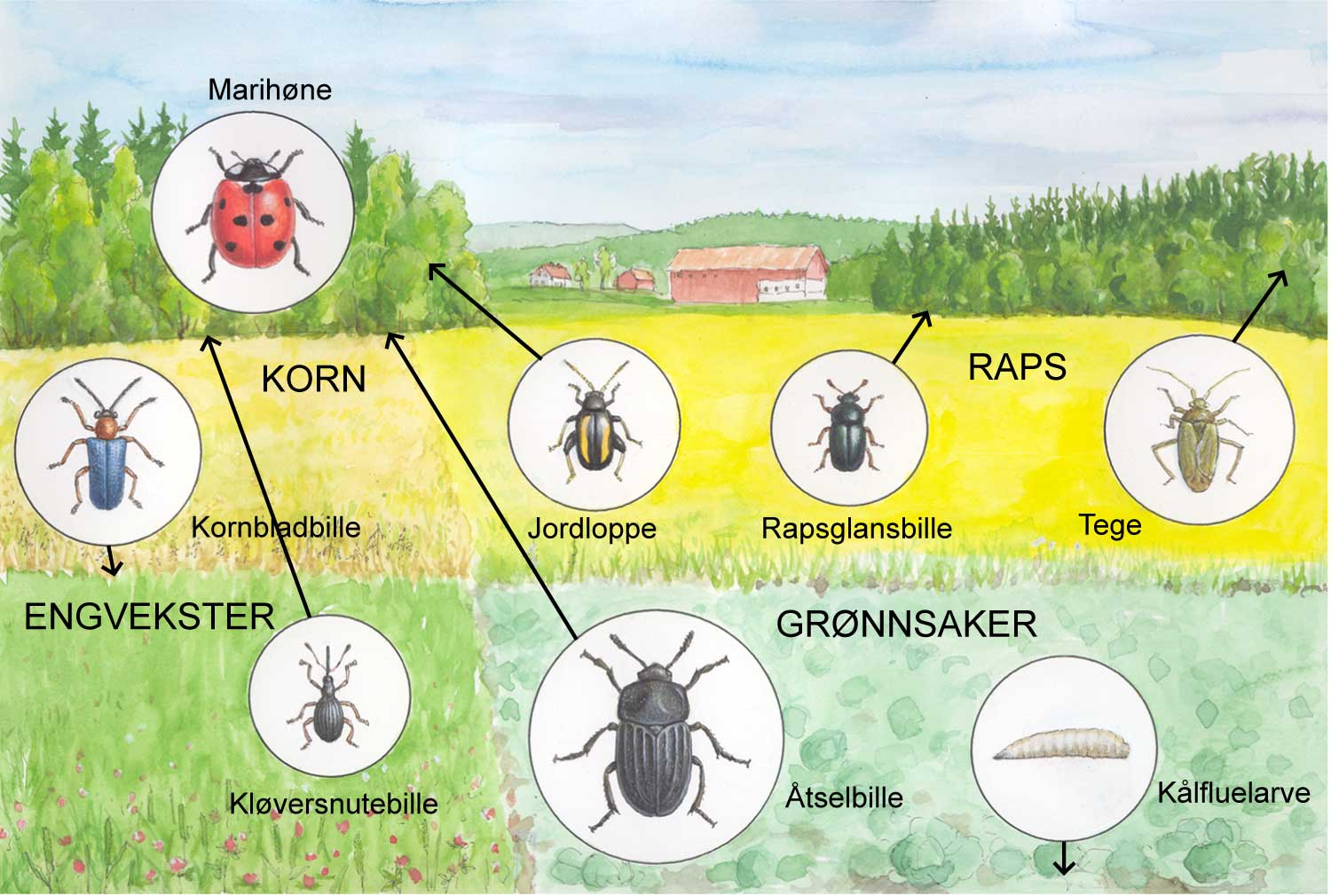
De fleste insekter flyr, og kan dermed aktivt søke seg til området hvor vertsplanten befinner seg. Noen er svake flyvere og blir fraktet med vind og luftmasser, men etterpå må også disse søke seg fram til de rette plantene. Denne søkeprosessen har flere trinn og forskjellige sanseorganer spiller inn. For å finne fram til riktig habitat (området hvor vertsplanten er) flyr insektet mot vinden og registrerer duftstoffene for å finne duften av vertsplanten. Når det nærmer seg planten overtar fargesansen og det velger å lande på den rette plantefargen, oftest mot en mørk bakgrunn (jord). Når det har landet bruker det smaksorganer for å bestemme om planten er av riktig slag. Hvis den er det, setter insektet i gang med å spise eller oppformere seg (figur 2.48). Hvis planten ikke er av det rette slaget forlater insektet planten og fortsetter søkingen. Oppformering skjer oftest som egglegging, men hos enkelte arter som bladlus kan det fødes levende unger («jomfrufødsel»).
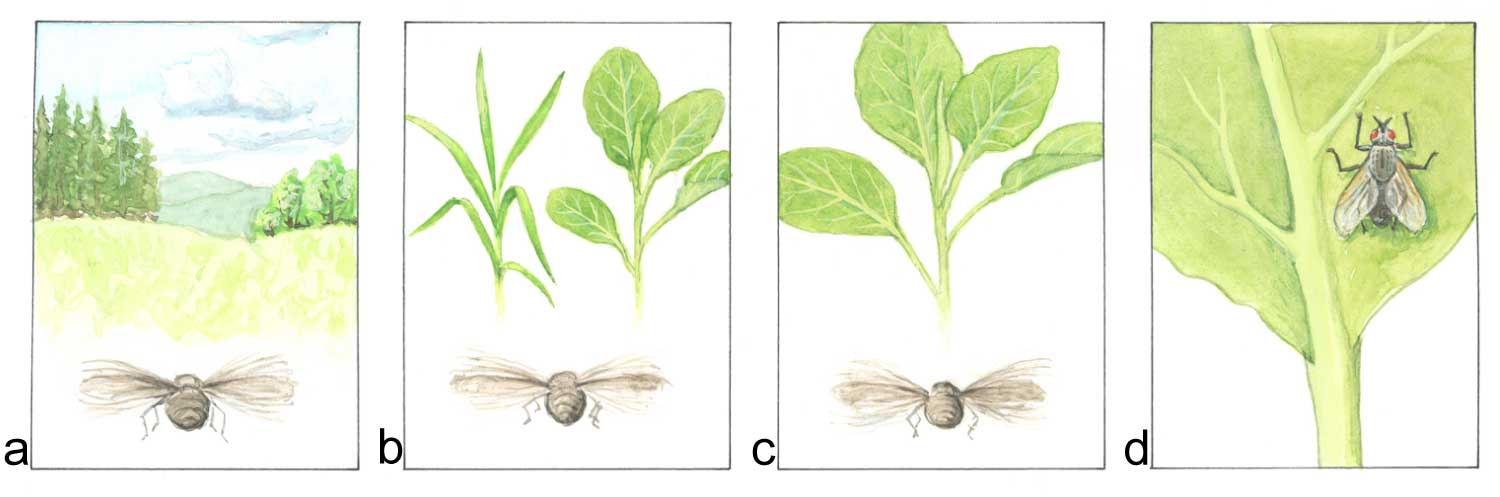
Insektfobi gjør at tilstedeværelse av småkryp i matproduktene er uønsket, også når de ikke gjør noe skade. I konvensjonelt landbruk forårsaker dette mye sprøyting som ikke har noe med vern av plantene å gjøre. En beslektet sak er «kosmetisk skade» på hagebruksprodukter. Det er viktig at forbrukerne veier sine krav på «ren mat» opp mot sine krav på skadefri og insektfri mat! Dette fører ofte til mye unødvendig bruk av ressurser for å tilfredsstille forbrukernes delvis ubegrunnede krav. I økologisk landbruk er det selvsagt uaktuelt å bruke giftstoffer i produksjonen.
Det er en kjent påstand at det er mindre skadedyrangrep i økologisk landbruk. Dette kan stemme, men årsakene kan være mange. Hvis vi tar som utgangspunkt at vekstskifte er helt vesentlig i økologisk landbruk, men mer tilfeldig og frivillig i andre driftsformer, kan dette være en viktig årsak til at vi kan oppleve mindre angrep, se kapittel .
I tillegg til vekstskifte er det som regel et større mangfold av vekster i økologisk landbruk enn i andre driftsformer. Det er mindre monokultur arealmessig, og dette gir dårligere vilkår for en plantespiser til å utvikle seg til å bli et skadedyr. Mat- og oppformerings-ressursene er redusert i omfang og dette begrenser utviklingen. I tillegg er det vanskeligere for et insekt som har kommet inn i området å finne fram til de rette vertsplantene når det er et større mangfold (figur 2.49). Dette skal vi se nærmere på i forbindelse med dekkekultur/ samplanting som er en bevisst bruk av mangfold i plantekulturen.
Rutinemessig bruk av plantevernmidler i andre driftsformer kan også være en årsak til mindre angrep i økologisk landbruk. Det er en kjent sak at de fleste kjemiske plantevernmidler slår ut nytteorganismer. Dette kan være både predatorer, parasitter og patogener. Se under avsnittet Biologisk kontroll (Kapittel 4) om betydningen av naturlig forekommende nytteorganismer. Det er vanlig at skadedyr utvikler resistens mot kjemiske midler raskere enn nytteorganismene gjør. Dette fører til en oppblomstring av skadedyrene fordi deres fiender fjernes.
En beslektet årsak er det som vi kaller for sekundære skadedyr. Det vil si plantespisende insekter som opprinnelig var til stede på våre kulturplanter uten å være så tallrike at de gjorde merkbar skade. En kjemisk behandling mot et annet insekt som var til stede i større mengder har drept nyttedyrene som var med på å holde den første arten i sjakk, og da blir et ikke-skadedyr til skadedyr.
På den negative siden er det lite å stille opp med i økologisk landbruk når et skadedyrangrep først er et faktum. Det er få ikke-kjemiske midler som er både tilgjengelige, effektive og godkjent for bruk i økologisk landbruk. Derfor er det meget viktig at angrep forebygges. Som nevnt er det viktigste forebyggende tiltak vekstskifte, men vi skal i de neste kapitlene også se på lokalisering av kulturen på gården, renhold i kulturen, jordarbeiding, så- og plantetid, sortsvalg og resistens mot skadedyr, bruk av dekkekultur/samplanting/fangstplanter og fysisk utestenging av skadedyr. Videre skal vi se på bruk av biologisk bekjempelse, d.v.s. bruken av nyttedyr og mikroorganismer. Og vi skal se på naturlige skadedyrmidler, som planteekstrakter, steinmel, m.m.
Agrios, G.N. 2005: Plant pathology -5thed.. Amsterdam, Elsvier, 922 s.
Andersson, S. og B. Eriksson 2001: Nematoder - världens vanligaste varelser. Faktablad om växtskydd, 56T, 8 s.
Conn, J.S. og R.E. Deck 1995: Seed viability and dormancy of 17 weed species after 9.7 years of burial in Alaska. Weed Science 43, s. 583-585
Danmarks JordbrugsForskning 2004: Ukrudtsbogen, 4. udgave.(Red. Madsen, K.H. og J. Jakobsen). Forskingscenter Flakkebjerg, Afdeling for Plantebeskyttelse, Flakkebjerg, Slagelse, 338 s.
Fykse; H. 1974: Studium av åkerdylle. II Utbreiing i Noreg, vokster og kvile – dels jamført med nærståande arter. Forskning og forsøk i landbruket Bind 25, s. 389-412
Fykse, H. og H. Sjursen 1992: Forelesingar i herbologi: I. Ugras Biologiske og økologiske eigenskaper, Landbruksbokhandleren, Ås-NLH, 108 s.
Fykse, H. og H. Sjursen 1992: Forelesingar i herbologi: II Rådgjerder mot ugras, Landbruksbokhandleren, Ås-NLH, 107s.
Grime, J.P. 1979: Plant strategies and vegetation processes. Chichester, Wiley, 222s.
Grime, J.P., J.G. Hodgson og R. Hunt 1988: Comparative plant ecology. A functional approach to common British species. Chapman og Hall, London, 741 s.
Hofsvang, T. og L. Sundheim 1990: Sjukdommer og skadedyr på jordbruksvekster. A/S Landbruksforlaget, 112 s.
Håkansson, S. 1995: Ogräs och odling på åker. Aktuelt från lantbruksuniversitetet 437/438 Mark Växter, Uppsala, 70 s.
Håkansson, S. 2003: Weeds and weed management on arable land - An ecological approach. CABI Publishing , UK, 274 s.
Kinzel, H. 1982: Planzen-ökologie und Mineral-stoffwechsel. Ulmer, Stuttgart, 534s.
Korsmo, E. 1935: Ugressfrø. Gyldendal Norske Forlag, Oslo, 175 s.
Korsmo, E. 1954: Ugras i nåtidens jordbruk. A/S Norsk Landbruks Forlag, Oslo, 635 s.
Lundkvist, A. og H. Fogelfors 2004: Ogräsreglering på åkermark. Rapport 6, Institutionen för ekologi och växtproduktionslära, Uppsala, 281 s.
Magnusson, C. og B. Hammeraas 1994: Potetcystenematodenes (PCN) biologi, smitteveier og bekjempelse. Faginfo 4, s.112-127
Palti, J. 1981: Cultural practices and infectious crop diseases. Advanced Series in Agricultural Science 9, Springer-Verlag, Berlin Heidelberg, 243 s.
Parry, D.W. 1990: Plant pathology in agriculture. Cambridge University Press, Cambridge, 385 s.
Radosevich, S.R. og J.S. Holt 1984: Weed ecology: implications for vegetation management. New York, Wiley, 265 s.
Rasmussen.I.A., S. Holm og S.I. Kristensen 1999: Plantebeskyttesle i økologisk jordbrug (Red. Rasmussen, I.A.) – En rapport fra en workshop, s. 1-84
Semb, K. 1996: Growth characteristics of spring barley and selected weeds .1. Effect of irradiance in growth chambers. Weed Research 36 (4), s. 339-352
Semb, K. 1996: Growth characteristics of spring barley and selected weeds .2. Effect of temperature and irradiance in growth chambers. Weed Research 36 (4), s. 353-367
Sjursen, H. 2001a: Change of the weed seed bank during the first complete six-course crop rotation after conversion from conventional to organic farming. Biological Agriculture and Horticulture 19, s. 71-90
Sjursen, H. 2001b: Biodiversitet av planter ved ulike driftsformer. Grønn Forskning 2/2001, s. 92-98
Sjursen, H. 2002: Ugrasutvikling ved økologisk grovforproduksjon. Grønn Forskning 2/2002, s. 216-222
Southwood, T.R.E. 1971: Farm management in Britain and its effect on animal populations. Proceedings Tall Timbers Conference on Ecol. animal control by habitat management, s. 29-51
Sundheim, L. 1982: Sjukdomar på korn og engvekster. Landbruksbokhandelen Ås–NLH, 132 s.
Teasdale, J.R. og C.S.T. Daughtry 1993: Weed suppression by live and desiccated hairy vetch (Vicia villosa). Weed Science 41, s. 207-212
Tischler, W. 1965: Agrarökologie. VEB Gustav Fisher Verlag, Jena, 499 s. Turner, S. J. 1996: Population decline of potato cyst nematodes (Globodera rostochiensis, G. pallida) in field soils in Northern Ireland. Ann. appl. Biol. 129, s. 315-322
Tømmerås, B.Å., T. Hofsvang, A. Jelmert, O.T. Sandlund, H. Sjursen og L. Sundheim 2003: Introduserte arter. Med fokus på problemarter for Norge. NINA (Norsk institutt for naturforskning) Oppdragsmelding 772, 58 s.
Whitehead, A.G. 1997: Plant nematode control. Cab International 1998, 384 s.
Øydvin, J. og B. Hammeraas 1973: Lethal low temperatures for the cyst content of the potato-root eelworms. Nematologica 19 (4), s. 568-569
Forebyggende tiltak for kontroll av skadegjørere er i tråd med grunntanken i økologisk landbruk, og har alltid spilt en sentral rolle innen slik dyrking. En god, helhetlig økologisk dyrkingspraksis vil bestå av et todelt aspekt: (1) Dyrkingspraksisen må ivareta kulturplantens behov, med tilstrekkelig næringsforsyning som spesielt viktig, men må samtidig (2) ikke legge forholdene til rette for oppformering og uakseptabel skade av ugras, sjukdomsorganismer og skadedyr. Solid kunnskap om kulturplanten og skadegjøreren hver for seg kreves for å få dette til, men også kunnskap om hvordan ulik dyrkingspraksis påvirker samspillet mellom kulturplante og skadegjører. I dette kapitlet vil derfor kunnskap om skadegjørernes livsstrategier, som vi har skrevet om i forrige kapittel, være et svært viktig grunnlag for tiltakene som beskrives.
Et gjennomtenkt vekstskifte er spesielt viktig i økologisk dyrking. Hvilke vekstskifter vi benytter, berører svært mange faktorer, ikke minst gjelder dette næringsforsyning og plantevern. I hvilken grad vekstskifte kan forhindre at plantevernproblem oppstår, er generelt avhengig av 3 sentrale egenskaper hos skadegjøreren:
Som utgangspunkt for dette kapitlet har vi benyttet en figur (figur 3.1) som viser sammenhengen mellom de 2 første av disse egenskapene hos en skadegjører, og hvilken gruppe tiltak som i størst grad kan kontrollere skadegjøreren.
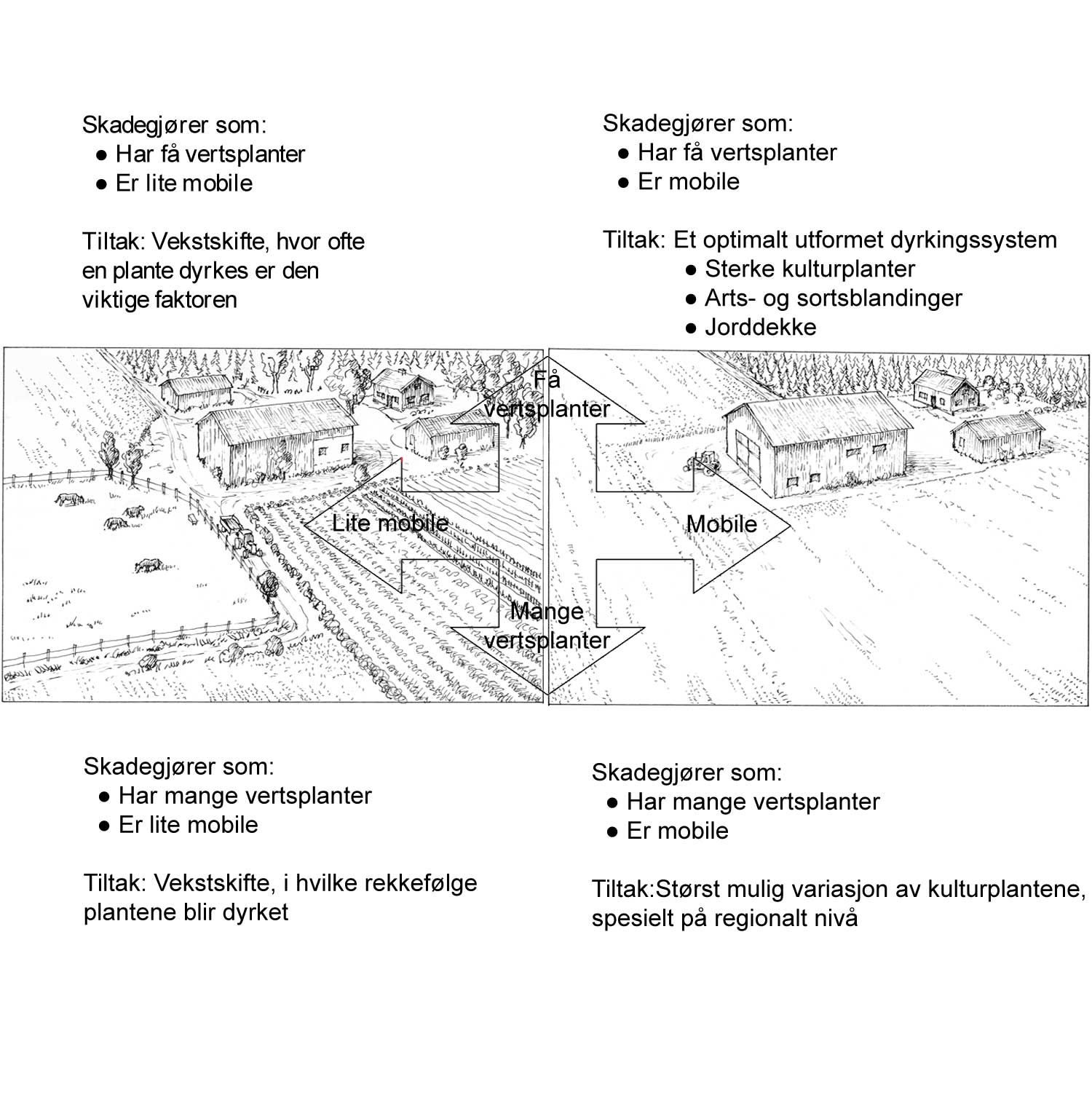
Som allerede nevnt, vil vi altså for mange skadegjørere finne aktuelle tiltak både innenfor vekstskifte og et optimalt utformet dyrkingssystem.
For sjukdomsorganismer og skadedyr er det ofte en svært spesifikk kobling mellom kulturplante og hvem som lever på og av den. En slik «ernæringsmessig kobling» har vi ikke mellom ugraset og kulturplantene, med unntak av snylteplanter, for eksempel timiansnyltetråd som lever på rødkløver og nærstående arter. Ugraset dukker opp i de kulturer hvor konkurranseforhold og ulike tiltak tilsier at de kan leve og produsere avkom. Enkelte ugras, for eksempel floghavre, har ganske bestemte krav til leveplass (kornåker) og vil bare unntaksvis skape problemer i andre kulturer. Som vi nevnte i forrige kapittel, vil sommerettårige frøugras, for eksempel meldestokk, hovedsakelig gjøre skade i kulturer der jorda bearbeides på våren, mens vinterettårige ugras også skaper problemer i høstsådde vekster. Den virkelig store generalisten blant ugras i Norge er kveke. Denne arten finner vi nær sagt i alle kulturvekstene vi dyrker, og det er hovedsakelig jordarbeiding som kan sette den tilbake. En svakhet ved kveka er likevel at den krever god lystilgang for å trives. Vekster som dekker godt, virker derfor sterkt hemmende på veksten. Havre gir således mindre kveke enn hvete. Mange av de forebyggende tiltakene som vi skal komme inn på i dette kapitlet, har to funksjoner, den ene er å drepe ugraset, mens den andre er å forsinke framspiringen og utviklingen av ugraset slik at det blir størst mulig størrelsesforskjell mellom ugras og kulturplante. Denne forskjellen er svært viktig for selektiviteten (dreper ugraset men ikke kulturplantene) ved mekaniske og termiske (flamming) tiltak.
For kontroll av skadegjørere som har få vertsplanter og som beveger seg lite, vil et gjennomtenkt vekstskifte stå svært sentralt. Å la det gå flere år mellom hver gang en kulturvekst dyrkes på samme areal, er ofte tilstrekkelig til å holde smittenivået under skadelig nivå. Hvor lang tid det bør gå før man kommer igjen med samme vekst, er avhengig av skadegjørerens oppformeringsgrad når vertsplante er til stede, og hvor raskt smittenivået synker ved fravær av vertsplante. Som vi allerede har vært inne på, vil bruk av resistente/tolerante sorter ofte kunne forsterke effekten av vekstskifte. Som et eksempel på dette kan det nevnes at hvis man har smitte av potetcystenematoder på gården sin, og skal dyrke mottakelige potetsorter, anbefales generelt et 6-årig vekstskifte. Hvis man benytter et omløp hvor man veksler mellom resistent og mottakelig potetsort, kan man gå ned på omløpstida og dyrke potet hvert tredje år.
For mange skadegjørere vil den mest effektive form for kontroll være at man hindrer smitte fra å komme inn på åkeren gjennom blant annet infisert jord og plantemateriale.
Å komponere et fornuftig vekstskifte handler imidlertid ikke bare om kontroll av en bestemt skadegjører. Bonden må tenke helhetlig, d.v.s ta hensyn til både skadegjørere, næringsforsyning og jordstruktur.
Vekstskifte er å dyrke ulike kulturplanter i en bestemt rekkefølge på et skifte. Tabell 3.1 viser eksempel på ensidig korndyrking kontra korndyrking med ert og kløvereng i omløpet.
Tabell 3.1 Eksempel på omløp med ensidig korndyrking og korndyrking med ert og kløvereng
| År 1 | År 2 | År 3 | År 4 | |
| Omløp 1: «Ensidig korndyrking» | Korn | Korn | Korn | Korn |
| Omløp 2: «Vekstskifte» | Kløvereng | Hvete | Havre + ert | Bygg m/gjenlegg |
Selv om «Omløp 2» er mer allsidig enn førstnevnte omløp, er heller ikke dette omløpet hva vi kan kalle et særlig «romslig» vekstskifte fordi det fremdeles er sterkt dominert av korn. Som vi skal høre mer om senere er det først når omløpet blir lengre, kanskje 6-8 år, og består av for eksempel radkulturer, flerårig eng og korn at man virkelig snakker om allsidig sammensatte omløp.
Samplanting har en del til felles med vekstskifte. Med samplanting menes ulike kulturvekster som står side om side på samme skifte (man kan kalle slik samplanting et «romlig» vekstskifte, i motsetning til et vekstskifte i tid som dette kapitlet omhandler). Fra småhagene kan vi hente et eksempel hvor løk og gulrot dyrkes i et «annenhver rad-system» spesielt med det for øyet å holde skadedyr borte.
Historiske kilder forteller oss at en dyrkingspraksis med vekstskifte har vært brukt svært lenge. For eksempel er det nevnt i litteraturen at vekstskifte var brukt under Han-dynastiet i Kina for mer enn 3000 år siden.
Utover 1900-tallet ble det i det moderne og mer industrielle landbruket en mer og mer gjengs oppfatning at eksterne innsatsfaktorer som kunstgjødsel og sprøytemiddel kunne erstatte vekstskifte uten at avlingene gikk ned. Vurderer vi bare ut fra et avlingsperspektiv kan vi vel også si at denne overgangen til mer ensidig plantedyrking mange steder lyktes ganske bra. Avlingsnivået i det «industrielle landbruket» er det jo ofte lite å si på, men det er også rapportert fra mange steder at avlingsnivået går ned etter flere års ensidig produksjon. Som nevnt tidligere, medførte denne type produksjonssystem at man også fikk med noen «uønskede blindpassasjerer». En annen, indirekte årsak til mindre bruk av vekstskifte, var overgangen til et mer mekanisert landbruk, noe som medførte at hesten som trekkdyr ofte forsvant fra gårdene, og bonden trengte dermed ikke å bruke noe areal til å produsere fôr til trekkrafta på gården.
Valg av vekstskifte påvirker mange faktorer som er viktig for avlingsnivået (tabell 3.2), som blant annet jordas innhold av organisk materiale, næringsforsyning og ulike skadegjørere.
Tabell 3.2 Faktorer som blir påvirket gjennom valg av vekstskifte
| Faktor | |
| Næringsforsyning: | Nitrogen |
| Fosfor | |
| Kalium | |
| Organisk materiale i jorda | |
| Jordstruktur: | Aggregatdannelse og stabilitet |
| Jordtetthet | |
| Vanninnfiltrasjonsevne | |
| Vannlagringsevne | |
| Skadegjørere: | Ugras |
| Sjukdomsorganismer | |
| Skadedyr |
Av disse er det nok næringsforsyning, særlig nitrogen, og å forebygge høye populasjoner/ bestander av skadegjørere som har blitt vektlagt mest når man har planlagt ulike omløp.
Næringsforsyning
Det er spesielt forholdet mellom nærende (inntektsside) og tærende (kostnadsside) vekster i et omløp som har blitt vektlagt. På kostnadssiden vil produksjon av matnyttige produkter som korn og grønnsaker hvor avlingene i større eller mindre grad høstes og fjernes fra produksjonsstedet, veie tungt. Kostnadene medfører blant annet at næringsstoffer som nitrogen og kalium fjernes fra jorda, at jordstrukturen blir dårligere, og at problemene med flerårige ugras øker. På inntektssiden kommer den delen av vekstskiftet som tilfører systemet nødvendige egenskaper, for eksempel bedring av jordstruktur og redusert bestand av flerårige ugras. Tilførsel av næringsstoffer som nitrogen gjennom bruk av belgvekster, for eksempel bønner eller erter i renbestand eller som del av engblanding, er også en særdeles viktige inntektspost. Bruk av underkultur i korn, som er en ren inntekt, og bruk av fangvekster for å overføre næringsstoffer til neste års kulturvekst, er også viktige faktorer i dette regnestykket. For en bestemt gård vil det være viktig å finne den rette balansen mellom inntekts- og kostnadssiden ut fra de begrensninger og muligheter som finnes i produksjonen. Kostnadsbalansen varierer sterkt mellom ulike situasjoner, blant annet vil klima og jordtype være av stor betydning.
«Næringsstoffer eller plantevern? Ja takk, begge deler»
For en økologisk bonde vil det ofte ikke være snakk om at det ene prioriteres framfor det andre, både næringsforsyning, plantevern og andre faktorer må ligge til grunn for valg av et bestemt omløp. Et godt vekstskifte vil være et system som produserer tilfredsstillende mengder av produkter av ønsket kvalitet, ut fra gitte begrensninger. Slike begrensninger kan være klima, jordart, tilgang på husdyrgjødsel, dyrkingsregler og tilgang på arbeidskraft. Systemet må også ha mest mulig stabile avlinger over tid. Økobonden erfarer ofte at ønsket om næringsforsyning og ugraskontroll kan stå i motsetningsforhold til hverandre. En underkultur som står utover høsten og samler nitrogen, vil for eksempel umuliggjøre stubbarbeiding mot kveke.
Hvilke vekster skal inn i omløpet og hvilken rekkefølge skal de ha?
Ved valg av vekstskifte er det viktig å finne ut hvilke vekster som skal utgjøre et omløp, og i hvilke rekkefølge de skal komme. Ut fra et næringsforsyningsaspekt vil det ofte falle naturlig at næringskrevende kulturer, som kanskje også krever god jordstruktur, for eksempel en del grønnsakskulturer, kommer rett etter eng eller et grønngjødslingsår (belgvekster i renbestand eller i blanding med grasart). Mer fordringsfulle kornarter, for eksempel hvete, vil på samme måte ofte komme etter kulturer som har gunstig ettervirkning på næringsstoffer og jordstruktur.
Hvilke typer skadegjørere kan man regulere ved hjelp av vekstskifte?
Det er skadegjørere med et begrenset vertsplantespekter, som er lite mobile og som ikke klarer å spre seg (smitte) ved fravær av vertsplante, som kan kontrolleres mest effektivt ved hjelp av vekstskifte. De skadegjørere som faller innenfor disse tre kriteriene er i all hovedsak jordlevende organismer av skadedyr og sjukdommer. Potetcystenematode og klumprot er typiske representanter for denne gruppen. Smittenivået i jorda faller imidlertid saktere ved fravær av vertsplante for klumprot enn hva tilfelle er for potetcystenematoder. Et annet aspekt ved vekstskifte og plantevern er at kulturvekster som ikke er vertsplanter, faktisk kan påvirke populasjonsutviklingen direkte hos ulike skadegjørere. Dette kan skje enten ved at de virker konserverende, for eksempel eng på klumprot, eller sanerende, for eksempel løkvekster som «lurer» larvene til potetcystenematoden til å klekke uten at vertsplante er tilgjengelig (tekstboks 3.1).
Når man planlegger hvilke kulturvekster som skal inn i et omløp, er det tradisjon for at man tenker mest på de vanlige vekstskiftefaktorene: Forholdet til vertsplanter, skadegjørerens mobilitet og hvor raskt smittenivået går ned ved fravær av vertsplante. Forsøk har imidlertid vist at det i en del tilfeller heller ikke er uten betydning hvilke «ikke-vertsplanter» som tas inn i et omløp. For eksempel har det i norske forsøk kommet fram at eng konserverer klumprotsmitte. Dersom en ønsker at smittenivået av denne skadegjøreren skal gå raskt ned, bør man altså ha lite eng i omløpet. De norske undersøkelsene har dessuten vist at pløying, sammenlignet med redusert jordarbeiding, gjør at smittenivået synker saktere. Det at eng konserverer smittenivået av en skadegjører har vi også sett i forsøk med potetcystenematoder. Årsaken til disse effektene av eng og pløying er litt uklare. At sporer og andre spredningsenheter havner dypt ned i jorda gjennom pløying, gjør muligens at de er vernet mot stor biologisk aktivitet. Ulike organismer i jorda kan for eksempel tenkes å bruke klumprotsporer som føde.
Det er også eksempler på at noen ikke-vertsplanter er spesielt gunstige for reduksjon av smittenivå for enkelte skadegjørere. Norske forsøk har blant annet vist at spesielt løkvekster, men også gulrot, er gunstige i et omløp for å senke potetcystenematodens smittenivå raskest mulig (se figur 3.2).
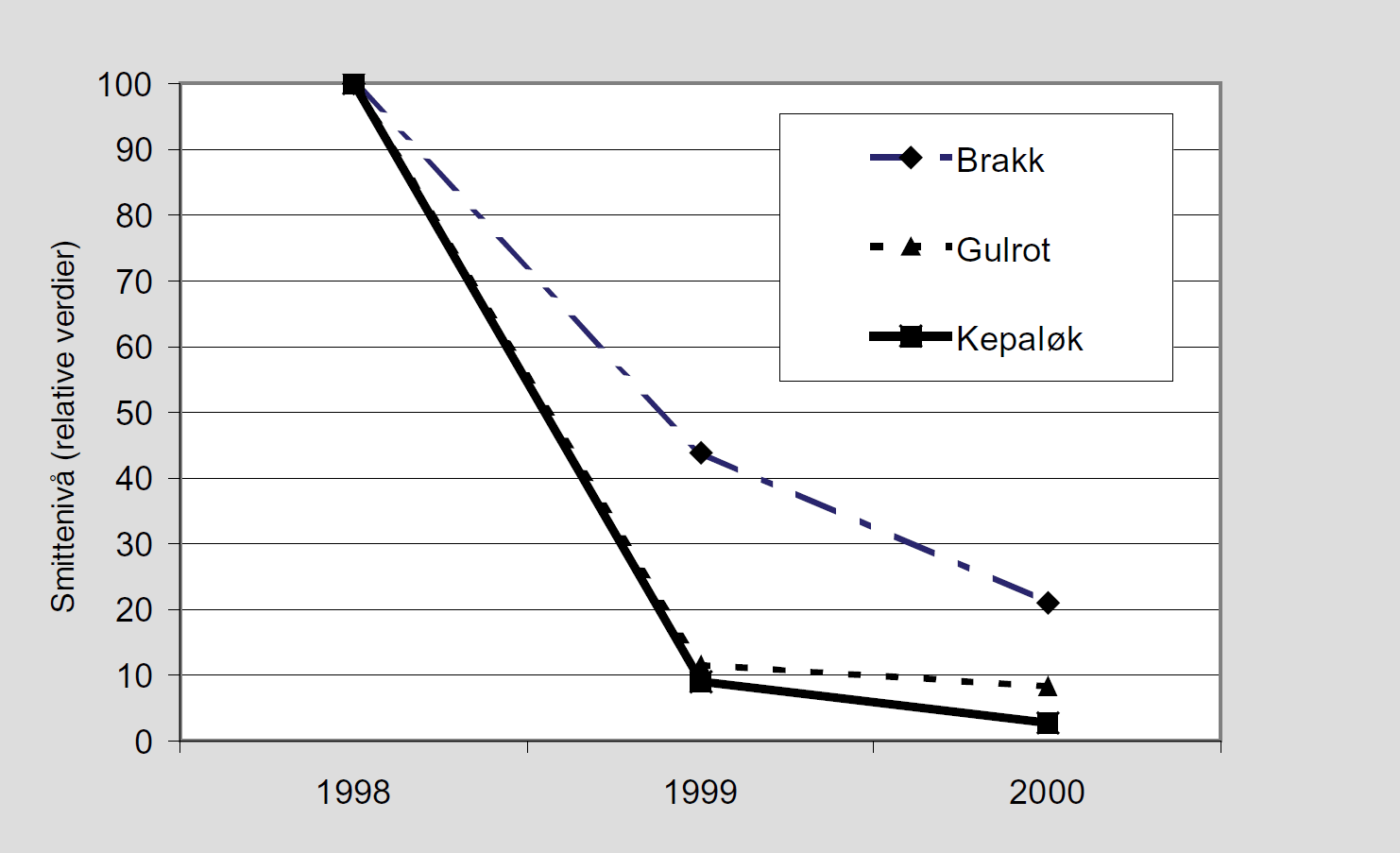
Innholdet av belgvekster blir derfor ofte mye større på økologiske gårder enn på gårder med konvensjonell drift. Generelt vet vi at ensidig dyrking av samme vekst ofte medfører at jordlevende skadegjørere oppformeres. For å motvirke dette kan man veksle mellom ulike belgvekster, for eksempel kløver, ert og åkerbønne. Hvis belgvekstene som integreres i et omløp, angripes av samme skadegjørere hjelper jo ikke dette. Det er derfor viktig å vite hvilke vertsplanteregister skadegjørerne har. Antakelig vil formering av planteparasittære nematoder (tekstboks 3.2) og sjukdomsorganismer (tekstboks 3.3) være det man kan frykte i denne sammenhengen. Det er derfor viktig å forebygge en utvikling som kan true den viktige «motoren» som belgvekster er, i økologisk dyrking. På den annen side vet man at bruk av grønngjødsling (innblanding av plantemateriale i jord) kan ha en renoverende effekt på mange skadegjørere.
Gjennom sin nitrogenfiksering er kløver og andre belgvekster svært viktige vekster i økologiske omløp. Flere nematodearter kan imidlertid redusere nitrogenfiksering fordi de skader belgvekstenes rotsystem. Graden av skade er avhengig av flere forhold, slik som nematodeslekt og -art, hvor mange nematoder som er i jorda, og vertsplantens mottagelighet. Angrep av nematoder kan redusere både avlinga, vekstenes overvintringsevne og levetid. Skaden nematodene gjør, kan forverres ytterligere ved at også andre Plantevern og plantehelse i økologisk landbruk organismer som sopp, bakterier og virus angriper de samme plantene. Dessuten kan belgvekstene oppformere nematodearter som også skader vekster som kommer senere i omløpet. Det kan medføre at vekster som dyrkes etter, eller sammen med belgvekster også kan få nedsatt avling hvis nematodene har denne veksten som vertsplante.
Viktige nematoder i belgvekster
De siste 40 årene er det ikke gjort systematisk kartlegging av utbredelse av nematoder som er skadegjørere på belgvekster i Norge. Her nevnes de viktigste slektene og artene av skadelige nematoder.
Kløvercystenematoden angriper og skader både hvitkløver og rødkløver. Særlig rødkløver er kjent å være svært mottagelig. Skadeterskelen ligger på ett egg per ml jord. I hvitkløver er det kjent at angrep av kløvercystenematoden påvirker utviklingen av Rhizobium-bakteriene slik at nitrogenfiksering blir redusert. Symptom på angrep kan sees som flekkvis dårlig vekst med små planter, i verste fall plantedød.
Stengelnematoden er kjent som et viktig nematodeproblem i rødkløver. Fire individer per 100 ml jord er nok til å gi betydelige skader. Angrepet sees lettest om våren eller forsommeren, og i gjenveksten på sensommeren. Symptomer kan være runde flekker eller store, sammenhengende flater med dårlig vekst i kløverfeltet. Angrepne planter blir korte med mange skudd hvor både basis, stengler og bladskaft kan bli sterkt oppsvulmet og forvridde. Ofte dør angrepne planter. Symptomene er de samme for rødkløver, hvitkløver, alsikekløver, og luserne.
Rotgallnematoder og rotsårnematodene har svært mange vertsplanter, også belgvekster. Det er ikke gjort forskning omkring skadene disse nematodene forårsaker på kløvervekster under norske forhold.
Potetråtenematoden er i tillegg til skadegjører på blant annet potet og gulrot også en skadegjører på kløver. Nematoden er observert i råtne røtter av rødkløver, og forsøk viser at den også er årsak til utgang av kløver. Viktige og gode vertsplanter for potetråtenematoden er også grønnsaker som kepaløk, pastinakk, rødbeter og selleri, og flere ugras som småsyre, åkerdylle, åkermynte, åkersvinerot og åkertistel.
Stuntnematoden lever fritt i jorda, og i motsetning til tidligere nevnte nematoder, spiser de på røttene fra utsiden. Stuntnematoder skader dessuten en rekke andre planteslag, og kan særlig observeres som svært redusert vekst i korn og gras.
Bekjempelse
Riktig valg av planteart, sorter og vekstrekkefølge i et vekstskifte er generelle og viktige tiltak mot nematoder. Bruk av resistente sorter av belgvekster mot for eksempel stengelnematoder er en del av dette. Foredling og testing for resistens mot stengelnematoder i rødkløver var tidligere et viktig arbeid som reduserte de store skadene i betydelig grad. Siden slik foredling og testing ikke lenger gjøres, må det forventes at skadeomfanget, særlig i rødkløver, tar seg opp igjen.
Ugrasrenhold er et nødvendig tiltak mot nematoder. Å fjerne gode vertsplanter for nematoder vil kunne redusere smittepresset på kulturplantene. Dette gjelder for alle nevnte nematoder unntatt cystenematoder som har et mer begrenset spekter av vertsplanter. Det samme kan også gjelde stengelnematoder med sine raser som er tilpasset spesielle plantearter.
Det sikreste rådet som kan gis, er at man varierer bruken av belgvekster mest mulig. Som et supplement til de vanlige belgvekstartene bør arter som fôrvikke/lodnevikke, luserne, legesteinkløver m.fl. vurderes.
Belgvekstene kan angripes av en rekke forskjellige jordboende sjukdomsorganismer (tabell 3.3). Enkelte sopper kan angripe flere ulike belgvekster, i blant også andre kulturplanter. I Norden er det likevel bare noen få sjukdommer som har vært spesielt vanskelige når man har dyrket en belgvekst hyppig. De mest kjente eksemplene så langt har vært visnesyke i erter og rotråte i kløver. Dersom dyrkingssystemet endres, og man begynner å dyrke belgvekster i større grad, kan imidlertid situasjonen forandre seg. Økt innslag av belgvekster kan medføre at mindre problematiske sjukdommer tiltar.
Mengda smitte (inokulum) samt klimafaktorer bestemmer infeksjonen
Angrepsgrad avgjøres av hvor mye smitte som finnes i jord og planterester i kombinasjon med klimafaktorer som temperatur og fuktighet. Andre faktorer som påvirker sjukdomsutviklingen, er jordtype og eventuelt sjukdomshemmende egenskaper i jorda. Smittenivået i jorda er som kjent sterkt avhengig av hvor ofte en belgvekst har vært dyrket i omløpet. Sortsvalg kan også bety noe. Et viktig spørsmål er hvordan tidligere dyrking av de vanlige kløverartene påvirker muligheten til å dyrke andre belgvekstarter.
Erfaringer fra Sverige
Forsøk i Sverige viser at smittepresset på kløver er relativt høyt uansett hvilke omløp man har, og at det heller ikke er avgjørende om man dyrker økologisk eller ikke. På den annen side, jo mer belgvekster man har dyrket, jo mer har man sett at svake sjukdommer har tatt seg opp. Som tabell 3.3 viser, er det funnet mange ulike sjukdomsframkallende sopper i svensk jord.
Vertsplantespekteret er en nøkkelfaktor
Omløpets sanerende effekt bestemmes i stor grad av de ulike skadegjøreres muligheter til å oppformeres i ulike vekster. Visse patogene sopper har en bred vertsplantekrets, hvilket betyr at de vil oppformeres i flere ulike kulturplanter. Andre sopper har bare et begrenset vertsplantespekter av nært beslektede vertsplanter.
Fusarium fra kløver smitter både bønne og ert.
Som nevnt, har flere belgvekstpatogener en bred vertsplantekrets, og svenske undersøkelser har vist at enkelte sopper kan gi sterke angrep på ulike belgvekstarter. For eksempel viste det seg at flere Fusariumarter fra rødkløver var sterkt patogene på blant annet ert og åkerbønne.
Visnesyke i erter, bare et problem i ert?
Visnesyke (Aphanomyces euteiches) er en sjukdom som først og fremst asossieres med ertedyrking. Et generelt råd er at man ikke bør dyrke erter oftere enn hvert sjette år for å hindre store problemer med denne soppen. I USA har man funnet at soppen kan være patogen også på rødkløver og luserne. I svenske undersøkelser har man ikke funnet at visnesyke angriper kløver, men foruten i erter er soppen funnet i luserne, legesteinkløver, fôrvikke og phaseolus-bønne.
Veien videre
I en engelsk studie fra 1980-tallet sies det at belgvekster bør sees på som «en og samme art» mht. jordboende sjukdommer. Forhåpentligvis er dette å trekke det litt for langt, men det kan slås fast at vi trenger mer kunnskap om disse soppenes vertsplantespekter her i landet.
Tabell 3.3 Kjente, helt eller delvis jordboende, plantesjukdommer på belgvekster i Sverige
| Belgvekst | Sjukdom | Sjukdomsorganisme |
| Phaseolusbønne | Spireskade | Pythium spp./Rhizoctonia solani |
| Svartråte | Thielaviopsis basicola | |
| Rotråte/spireskade | Fusarium spp. | |
| Luserne | Phoma råte | Phoma medicaginis |
| Visnesyke | Verticillium dahliae | |
| Rødkløver | Rotråte/spireskade | Fusarium spp. |
| Kløverråte | Sclerotinia trifoliorum | |
| Spireskade | Pythium spp. | |
| Hvitkløver | Spireskade | Pythium spp. |
| Rotråte/spireskade | Fusarium spp. | |
| Stolon- og rotråte | Cylindrocarpon spp. | |
| Phoma sp . | ||
| Colletotrichum spp . | ||
| Åkerbønne | Sjokoladeflekksyke | Botrytis cinerea/ B. fabae |
| Rotråte/spireskade | Fusarium spp. | |
| Rhizoctonia spp . | ||
| Pythium spp . | ||
| Phoma spp. | ||
| Cylindrocarpon sp . | ||
| Ert | Visnesyke | Aphanomyces euteiches |
| Svartråte | Thielaviopsis basicola | |
| Spireskade | Pythium spp. | |
| Rhizoctonia solani | ||
| Rotråte/spireskade | Fusarium spp. | |
| Stengelråte | Sclerotinia spp. | |
| Stengel og rotråte | Phoma medicaginis var. pinodella |
Etter Levenfors, Lager og Gerhardson 2001
Vekstskifteeffekten på ugraset kan vi si er en «puslespilleeffekt» ved at vi setter sammen brikker med ulike kulturvekster og jordarbeidingsmåter. I tillegg til disse brikkene med kulturer og jordarbeiding vil dessuten mange andre faktorer som klima, jordtype og gjødsling være bestemmende for både sammensetning og mengde ugras.
Ugraseffekten av vekstskifte kan vi derfor si er en langtidseffekt av hvordan ugraset blir påvirket av kulturplantene vi dyrker, inkludert effekten av jordarbeiding som gjennomføres:
For noen kan det kanskje virke litt rart at effekten av jordarbeiding inkluderes som en del av vekstskifte, men valg av kulturvekst og gjennomføring av jordarbeiding er så nært knyttet til hverandre at det er svært vanskelig å skille dem fra hverandre.
Kombinasjoner av at ugras, først og fremst flerårige arter, først tynes gjennom jordarbeiding for så å utsettes for konkurranse fra en kulturvekst, er også interessant. Et godt eksempel er kombinasjonen av utsulting av kvekejordstengler ved skålharving etterfulgt av dyp nedpløying, med etablering av en kultur med god konkurranseevne. Slike kombinasjoner av jordarbeiding og konkurranse er svært aktuelt for kontroll av alle flerårige ugras ved økologisk dyrking (dette belyses mer under avsnittet om grønngjødsling).
Det finnes ikke spesielle vekstskifteomløp som i seg selv, og uten bruk av effektive direkte tiltak, hindrer alle typer ugras i å spire og etablere seg. Dette skyldes at alle ugras innenfor de ulike biologiske ugrasgruppene (sommerettårige, vinterettårige m.fl.) enten har klare fordeler eller begrensninger hva gjelder sitt nærvær i forskjellige kulturvekster. Effekter på flerårige ugras er antakelig viktigere enn virkningen på frøugras.
Hvilke ugras vokser i de ulike kulturer?
I et omløp som er dominert av kulturvekster som framelsker bestemte ugras, eller en gruppe av ugras, og hvor man ikke setter inn effektive tiltak, vil ugraset etter hvert bli svært tallrikt. Eksempler på dette kan vi hente fra tabell 2.1. Ugras eller ugrasgrupper som er merket med ++ eller +++ for de forskjellige kulturvekster, har potensiale til å øke sin utbredelse sterkt hvis man dyrker disse for ensidig.
Noen eksempler kan nevnes:
Ulike kulturvekster har svært forskjellig potensiale for å konkurrere med ugraset. Vekster som dekker jorda tidlig, og som har høy tørrstoffproduksjon, har generelt best evne til å konkurrere med ugraset. Eksempler på slike vekster er korn og oljevekster. En veletablert og tett eng er et annet eksempel på en kulturvekst som både er sterk mot ettårige og mange flerårige ugras. En vekst som poteter etablerer et tett bestand relativt sent om sommeren, men vil etter den tid konkurrere bra med ugraset.
Det er ingen lett oppgave å sette opp en fasit på hvordan ulike vekstrekkefølger vil påvirke ugrasutviklingen. På den annen side kan man lett definere hvordan problemer best mulig skal forebygges (og det gjelder egentlig ikke bare ugras, men alle skadegjørere):
En størst mulig variasjon i omløpet vil hindre at problemer bygger seg opp etter hvert. Denne variasjonen vil omfatte hvordan plantene rent morfologisk ser ut, hvilke mønster de blir sådd eller plantet i, så- og plantetidspunkt, hvilke direkte tiltak som benyttes med mer.
Vi vet at det, antakelig uten unntak, finnes ugras som har en livsstrategi som alltid passer de ulike kulturplantene. For korn er det kanskje spesielt vanskelig. Ikke bare har vi floghavren (som også er en kornart), som selvfølgelig trives under de samme forhold som «kulturkornet». Arter som åkertistel, åkerdylle og kveke trives også svært godt under de samme forhold som kornet. Dette tilsier at problemene vil øke med tida hvis vi prøver å dyrke korn ensidig over tid. Økobondens utfordring vil derfor være å sette opp slike vekstfølger at de ulike enkeltarter ikke får sjanse til å danne bestand som truer en fornuftig avlingsstabilitet på gården. Også mindre forandringer i et omløp, for eksempel ved at en kornbonde dyrker høstkorn i tillegg til vårkorn, gir små, men viktige bidrag til å få et mer robust system. Innlemmes også radkulturer i omløpet, vil bonden kunne gjennomføre radrensing som også vil hemme de tre nevnte ugrasartene.
Dette høres veldig greit ut, men økobonden vil selvfølgelig, som andre bønder, også måtte forholde seg til mange andre faktorer, som landbrukspolitikk (hvordan kanaliseringspolitikken påvirker vekstskiftemuligheter), økonomi og «antall timer i døgnet».
Vekstskiftevalg vil også dreie seg om å finne kulturer som muliggjør forskjellige mekaniske tiltak, for eksempel radrensing.
«Fordel for ugraset» kan vendes til «fordel for bonden»
Det er et problem for bonden at ensidig dyrking av bestemte kulturvekster favoriserer enkelte ugras, men trøsten er at dette «fortrinnet for ugraset» kan vendes til en motsatt situasjon: Når effektiv kontroll gjennomføres, kan den kulturplante som uten bekjempning er mest gagnlig for ugraset, endres til den veksten i omløpet som setter ugraset mest tilbake.
For å illustrere dette kan vi ta et eksempel fra grønnsakdyrkingen: De sommerettårige artene er avhengig av jordarbeiding om våren for å spire godt. Når grønnsakbonden om våren gjennomfører jordarbeiding før såing eller planting, blir dette kravet innfridd og flere frø spire. Hvis grønsakbonden så gjennomfører tiltak som dreper de oppkomne ugrasplantene, og dermed hindrer ny produksjon av frø, vil han redusere frøbanken mer i slike kulturer enn i andre.
I praksis vil det nok ofte være slik at det i grønnsaker skjer en oppblomstring av frøugraset fordi for mange frøplanter får stå i fred og formere seg. Hvis derimot direktetiltakene (mekaniske og termiske tiltak supplert med luking) som gjennomføres er gode nok, vil altså frøbanken reduseres.
Det er imidlertid en stor forskjell mellom den økologiske og konvensjonelle bonden når det gjelder muligheten til å utnytte at gitte ugras kommer sterkt i visse kulturer. Den konvensjonelle bonden tar ofte fram åkersprøyta i denne situasjonen og kan stort sett i enhver kultur sette inn et effektivt sprøytetiltak. Økologiske bønder har ikke denne muligheten. I radkulturer har de ulike former for ugrasregulering tilgjengelig, mens situasjonen er langt vanskeligere i andre kulturer. For eksempel vil man i korn med vanlig radavstand ha få muligheter til å gjøre noe med rotugras på større arealer.
Mange undersøkelser i utlandet har konkludert med at behandling av kulturvekstene i et omløp (effektiviteten i radrensingen, antall slåtter i enga osv.) er minst like avgjørende som valget av vekster i omløpet.
Noen eksempler
Eng: Frøugras
Forsøk bl.a. i Norge har vist at flere år med eng gir en markant minkende frøbank. Den økte imidlertid raskt igjen da det ble dyrket «åpne kulturer» som korn og ulike radkulturer (figur 2.19). Hvor mye frøbanken øker i åkerkulturene er selvfølgelig avhengig av hvor god kontroll en har på ugraset i disse kulturene.
Eng: Flerårig ugras
Langvarig eng vil begunstige engugras som for eksempel høymole. I åkervekster vil rotugras som åkertistel og åkerdylle dominere. Tabell 3.4 viser at hyppigheten av disse to artene er størst i vårkorn, betydelig lavere i høstkornet, og at de dessuten blir mindre hyppige i eldre eng.
Tabell 3.4 Forekomst av dyllearter og åkertistel i ulike kulturer/vokseplasser som prosent av undersøkte eiendommer (etter Fykse 1974)
| Kultur eller vokseplass | |||||||||
| Korn | Radkulturer | Gras/Eng | |||||||
| Vår- korn | Høst- korn | Grønn- saker |
Poteter og rot- vekster | Første årseng | Eldre kunsteng | Kultur- beite | Natur- eng | ||
| Flerårige arter: | Åkerdylle | 60 | 32 | 50 | 57 | 35 | 30 | 21 | 21 |
| Åkertistel | 61 | 33 | 50 | 57 | 42 | 47 | 39 | 39 | |
| Ettårige arter: | Haredylle | 43 | 23 | 40 | 39 | 12 | 7 | 7 | 4 |
| Stivdylle | 47 | 21 | 44 | 44 | 13 | 5 | 5 | 3 | |
Med unntak av tre fylker var åkertistel mer vanlig enn åkerdylle. I åpenåker stod de to artene svært likt, men med mer åkertistel i grasmark. Vårkorn inneholdt disse vekstene oftest, men poteter og rotvekster kom ikke langt bak. Haredylle og stivdylle fins mest i åpenåker, lite i grasmark.
Alle arter var mest vanlig på mold- og myrjord, men forekom også på leirjord, sand- og grusjord.
Korn: Floghavre og flerårig ugras
All erfaring tilsier at vi nærmest ber om problemer hvis vi prøver å dyrke økologisk korn i ensidige omløp. Vi har allerede vært inne på effekten av vanlig eng og grønngjødslingseng. Et annet aktuelt tiltak er å ta inn kulturer som kan radrenses. Bondens, veilederens og forskerens oppgaver i dag er derfor å finne en del «tålegrenser». En slik «tålegrense» forteller hvor mye korn vi kan ha i et omløp, uten at problemene med ugras vokser oss over hodet.
Et allsidig vekstskifte er svært viktig for å motvirke sjukdomsproblemer på planter. I hvor sterk grad sjukdommene kan motvirkes og hvilke sjukdommer man påvirker lettest, vil variere. Tidligere har vi beskrevet sjukdomssoppenes ulike måter å leve og overvintre på. Vi har sett at noen kan leve på en rekke ulike verter mens andre må ha en helt spesiell vert for å overleve. Soppens preferanser er avgjørende for hvordan et vekstskifte påvirker forekomsten av en sjukdomsorganisme. Sjukdommer som overlever og spres fra planterester i jord og samtidig har smalt vertsplantespekter, vil være de som lettest bekjempes ved hjelp av vekstskifte. Tørråte er en organisme med smalt/spesifikt vertsplantespekter (potet/tomat), men med stor mobilitet/spredningsevne (spres svært raskt både innen en åker og over større avstander). Når soppen tar luftveien til hjelp, vil ikke vekstskifte ha samme effekt som når soppsporer spres fra gamle planterester i samme åker. Plassering av de ulike kulturene på ulike skifter vil likevel kunne ha betydning for denne typen skadegjørere (kan sees på som «vekstskifte i rom» kontra «vekstskifte i tid»). Tidligpotet bør for eksempel ikke dyrkes ved siden av sene poteter. Tørråte vil kunne oppformere seg i tidligpotetåkeren og produsere mye ny smitte til de senere potetene som vil ta mer skade av infeksjon med tørråte.
Mjøldogg i hvete eller bygg spres med sporer med vind og vil som oftest finne veien til sine verter selv ved et godt vekstskifte. Her gjelder imidlertid det samme prinsippet om oppformering av smitte. Plassering av høsthvete i nærheten av vårhvete vil gi tidligere mjøldoggsmitte i åkeren med vårhvete og dermed større skade.
Som nevnt vil en sjukdom som overlever på planterester i jorda, i stor grad påvirkes av vekstskifte dersom de ikke er altfor bredspektret med hensyn til vertsplanter. Smitte av sjukdommer som byggbrunflekk, havrebladseptoria eller grå øyeflekk vil reduseres betydelig i løpet av bare en sesong med en annen kornart eller annen vekst. En sjukdom som storknolla råtesopp vil derimot kreve opptil 6 år med en annen kulturvekst (ikke oljevekster eller erter/bønner). Dette skyldes at den overlever i form av sklerotier med svært lang levetid i jorda. Klumprot i kålvekster og visnesyke i erter er også eksempler på sjukdommer som kan leve lenge i jorda.
Et godt vekstskifte alene er sjelden tilstrekkelig for å redusere forekomsten av patogener som har overlevelsesorganer som nevnt over, eller som har et bredt vertspekter og altså kan angripe mange av kulturvekstene. Imidlertid vil en kombinasjon av et godt vekstskifte og god jordarbeiding, tilførsel av organisk materiale/gjødsel (grønngjødsel, husdyrgjødsel), god vannhusholdning og god ugraskontroll kunne være suksessfullt også overfor slike vanskelige skadegjørere.
Det finnes gode eksempler fra utlandet på at vanskelige, jordboende sjukdommer i blant annet jordnøtter og bomull kontrolleres ved hjelp av et godt planlagt vekstskifte sammen med tiltak som listet over. Et godt planlagt vekstskifte vil i mange tilfeller kunne bremse oppbygning av sjukdomssmitte i jorda, også av de mer problematiske sjukdommene.
En populasjon av et skadedyr som finner en gunstig vertsplante på feltet, vil kunne bygge seg opp år etter år. Denne utviklingen kan brytes ved vekstskifte og dyrking i ett eller flere år av en ikke-vertsplante.
Vi skal diskutere litt mer inngående, i relasjon til skadedyr, de tidligere nevnte hovedpunktene vedrørende biologien til en skadegjører:
I noen tilfeller med vekstskifte, ikke minst på små arealer, er det viktig å ta hensyn til de kulturene naboen dyrker. For skadedyr med 1-årig livssyklus, for eksempel gulrotflue, vil ett år uten gulrot være nok til å bli kvitt puppene i jorda. Hvis derimot gulrotfelter finnes i nærliggende områder, vil voksne gulrotfluer fly inn fra omgivelsene og kolonisere feltet på nytt.
I årene 1891–1939 var smellerlarver (kjølmark) det oftest omtalte skadedyret i korn. Billene legger egg i gammel eng, og etter pløying går larvene over i etterfølgende kultur. En vesentlig årsak til de omfattende angrepene i korn var derfor gammel eng som forkultur. Rotvekster og potet er særlig utsatt for angrep av kjølmark, og bør derfor ikke dyrkes før minst ett år etter eng eller korn, der man vet at det har vært kjølmarkangrep.
Utvidet dyrking av en kulturplante innen et distrikt kan gi stor økning i skadedyrpopulasjonene. Fra slutten av 1940-årene økte dyrkingen av løk sterkt på Toten, senere også i Vestfold. Dette førte til omfattende angrep av løkflue utover i 1950-årene inntil en fikk effektive metoder for bekjempelse.
Enkelte ganger kan vekstskifte med hell omfatte hele distrikter for å få bukt med farlige skadedyr. I Grimstad-distriktet ble det i 1940-årene innført karanteneordning med forbud mot dyrking av erter i bestemte områder på grunn av ertegallmygg. Systemet med vekstskifte basert på distrikt praktiseres i konservesert i dag. Til tross for utstrakt dyrking innen regioner, har en unngått angrep av for eksempel ertevikler som tidligere var et alvorlig skadedyr i ertedyrkingsområdene. Derimot har intensiv dyrking av såkalte «Ringerikserter» i Hole i Buskerud de siste årene ført til en opphopning av populasjonen av ertevikler. Et annet eksempel på slikt regionalt vekstskifte er i Rogaland hvor gulrot og andre skjermplanter ble dyrket i én dal et år og nabodalen det neste året. Et år uten skjermplanter var nok til å tømme området for gulrotfluer. Dette var et frivillig tiltak og kan være vanskelig å gjennomføre i dagens samfunn. Slikt regionalt vekstskifte har også vært foreslått i nyere tid i forbindelse med kålflue i korsblomstrede vekster.
En negativ følge av vekstskifte er at skadedyrenes naturlige fiender ikke får tid til å bygge seg opp før kulturen og verts- eller byttedyret forsvinner fra stedet. Men fordelene med vekstskifte er langt flere enn ulempene.
Grønngjødsling er et samlebegrep for ulike typer vekster som i utgangspunktet dyrkes for å tilføre jorda gjødsel, hovedsakelig nitrogen. Ulike belgvekstarter, med sin evne til å ta opp nitrogen fra lufta (nitrogenfiksering), er de vekstene man oftest forbinder med grønngjødsling. Andre ikke-nitrogenfikserende arter brukes også ofte, enten i blanding med belgvekstene, eller alene. Forekomsten av ugras kan både reduseres og tilta gjennom en periode med grønngjødsling. Et veletablert grønngjødslingsbestand av konkurransesterke arter som lodnevikke eller rødkløver kan effektivt redusere ugraset. På den annen side vil en grønngjødslingsart som slipper gjennom mye lys, og som samtidig kanskje også er dårlig etablert, føre til oppformering av både frøugras og flerårige arter.
Grønngjødslingsvekstene kan også ha flere andre interessante egenskaper, blant annet har det vært en god del fokus på arter med dypt rotsystem som har evna til å ta opp næringsstoffer fra store jorddyp. Disse artene har en «pumpevirkning» på næringsstoffene på den måten at de blir gjort tilgjengelige for den etterkommende veksten, for eksempel grønnsaker, når grønngjødslingsvekstene pløyes eller freses inn i jorda. Grønngjødslingsvekster med dyp pålerot, for eksempel legesteinkløver, er dessuten kjent for å ha en gunstig effekt på jordstrukturen.
Under våre klimaforhold er det enkelte sjukdommer som er spesielt besværlige i systemer med mye engbelgvekster. I en norsk undersøkelse av kløverrøtter i eldre eng, ble det funnet Fusarium i relativt mange prøver av kløverrøttene. En større undersøkelse i Sverige, som nettopp tar for seg forekomst av sjukdommer i belgvekster ved økologiske dyrkingssystemer, viste at den vanligste Fusariumarten på korn og gras i Norge (Fusarium avenaceum) også er vanlig forekommende på engbelgvekster og andre belgvekster der. Kløverråte forårsakes av en annen sopp som danner sklerotier. Disse sklerotiene kan man finne nede i rothalsen på syke planter. Begge disse sjukdommene smittes via jord. Grønngjødslingsvekster kan integreres i ulike omløp på svært mange måter (for muligheter i korn, se figur 3.3).
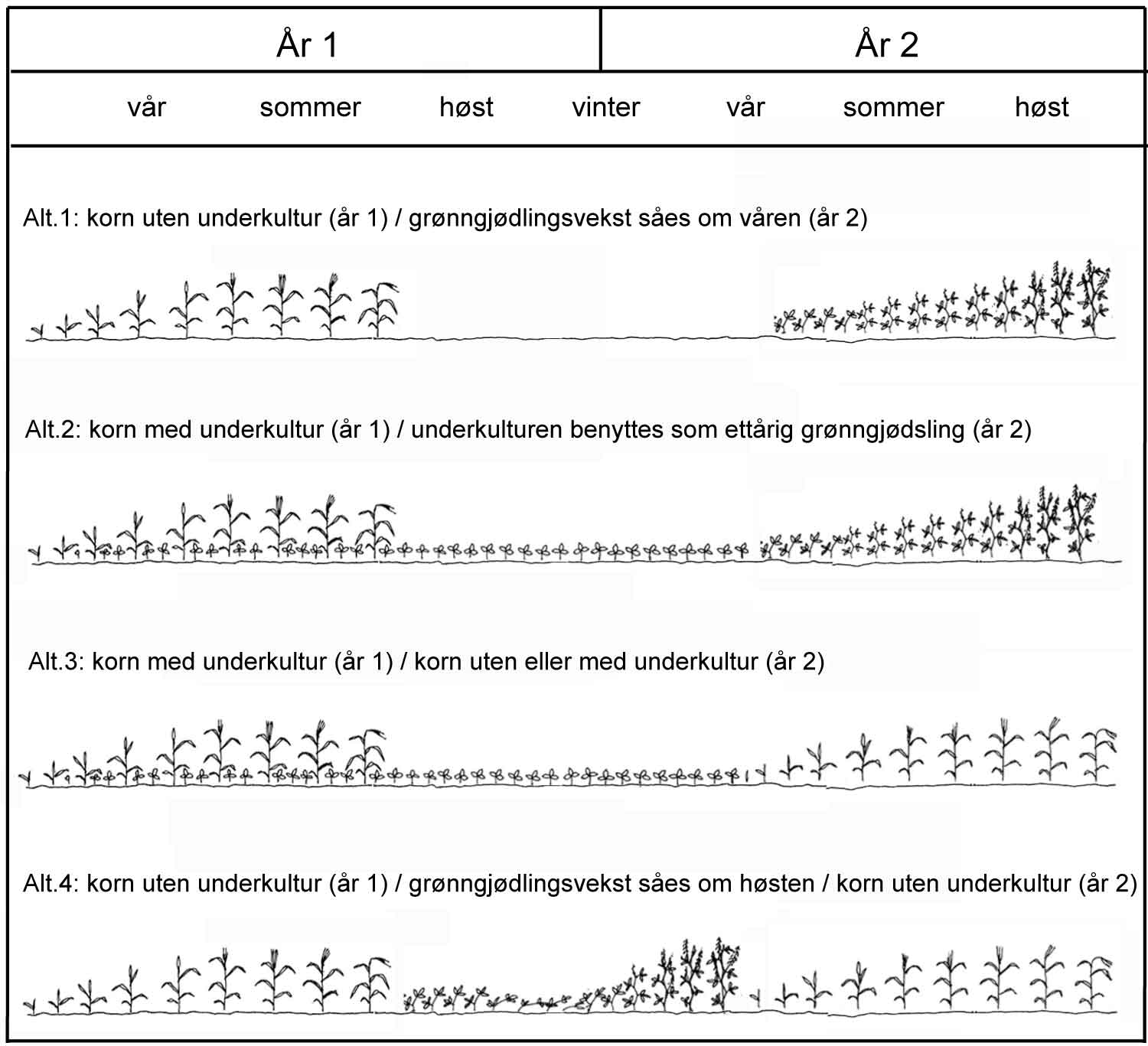
Se også figur 3.4 og tekstboks 3.5.
Frøene til mange ugras trenger en lysimpuls for å spire. Behovet for lys er en økologisk tilpassing for å hindre at frøet, og da spesielt arter med små frø, spirer i situasjoner hvor de svake spirene har små muligheter til å overleve. Frøets muligheter til å reagere på lys styres av fytokrom, et lysabsorberende pigmentsystem. Dette pigmentsystemet avhenger ikke bare av mengde lys, men også lyskvalitet. Ved f.eks. solens opp- og nedgang er det en høy andel mørkerødt lys, hvilket er mindre effektivt til å indusere frøspiring. To tyskere, Hartmann og Nezadal, var de første (i 1990) som foreslo at kunnskapen om frøenes lysbehov kunne utnyttes i praksis for å redusere antall ugrasfrø som spirer. Det har siden vært mange undersøkelser omkring slikt «mørkearbeide». Noen av disse er utført om natten, andre med tildekket jordarbeidingsredskap om dagen (se figur 3.22).
En gjennomgang av undersøkelsene viser at nedgangen i antall ugrasplanter varierer fra 60-70 % i de mest lovende til nesten ingen utslag i andre. Et toårig forsøk i Sverige viste dessuten stor variasjon, fra over 60 % reduksjon det ene året til bare 14 % reduksjon året etterpå. Den relativt dårlige overensstemmelsen mellom ulike undersøkelser kan bero på ulike forutsetninger (bl.a. lysnivåer, artssammensetning, bearbeidingsintensitet) som forsøkene er utført under. Effekten av jordarbeiding i mørke kan bero på to ulike forklaringer; 1. Reduksjon i antall spirte frø, og 2. Forsinket framspiring, som kan skyldes at frø nærmest jordoverflata (1-4 mm dybde) først etter å ha akkumulert tilstrekkelig av det svake lyset som trenger ned gjennom jorda, er i stand til å spire. Av den grunn kan også tidspunktet for registrering være avgjørende for resultatet og kanskje forklare en del av variasjonen mellom undersøkelser.
Både redusert og forsinket ugrasspiring er interessant fordi det styrker kulturveksten i konkurransen. Jordarbeiding i mørke vil likevel i de fleste tilfeller ikke være effektivt nok til at tiltaket alene er godt nok. Potensialet ligger derfor mer i å la «nattarbeid» inngå i en strategi sammen med andre tiltak, f.eks. ulike direkte tiltak. Jordarbeiding i mørke kan medføre økt selektivitet ved senere radrensing eller ugrasharving ved at kulturveksten får et forsprang i forhold til ugraset. På den annen side kan dette tiltaket, i likhet med andre kontrolltiltak, føre til at tolerante ugras selekteres, i dette tilfellet arter som spirer relativt uavhengig av lystilgangen. Dette tilsier at jordarbeiding i mørke bør integreres i en bekjempningsstrategi. Det er fremdeles uklart hvor mørkt det må være i forbindelse med jordarbeidinga for å unngå spireinitiering. Som en kuriositet i så henseende kan det nevnes at det i tyske forsøk har vært brukt infrarøde nattbriller for å kunne kjøre uten lys fra traktoren, mens det i danske forsøk, i hvert fall tidligere, har vært brukt framlys på traktoren uten at man har ansett det for å kunne påvirke resultatene.
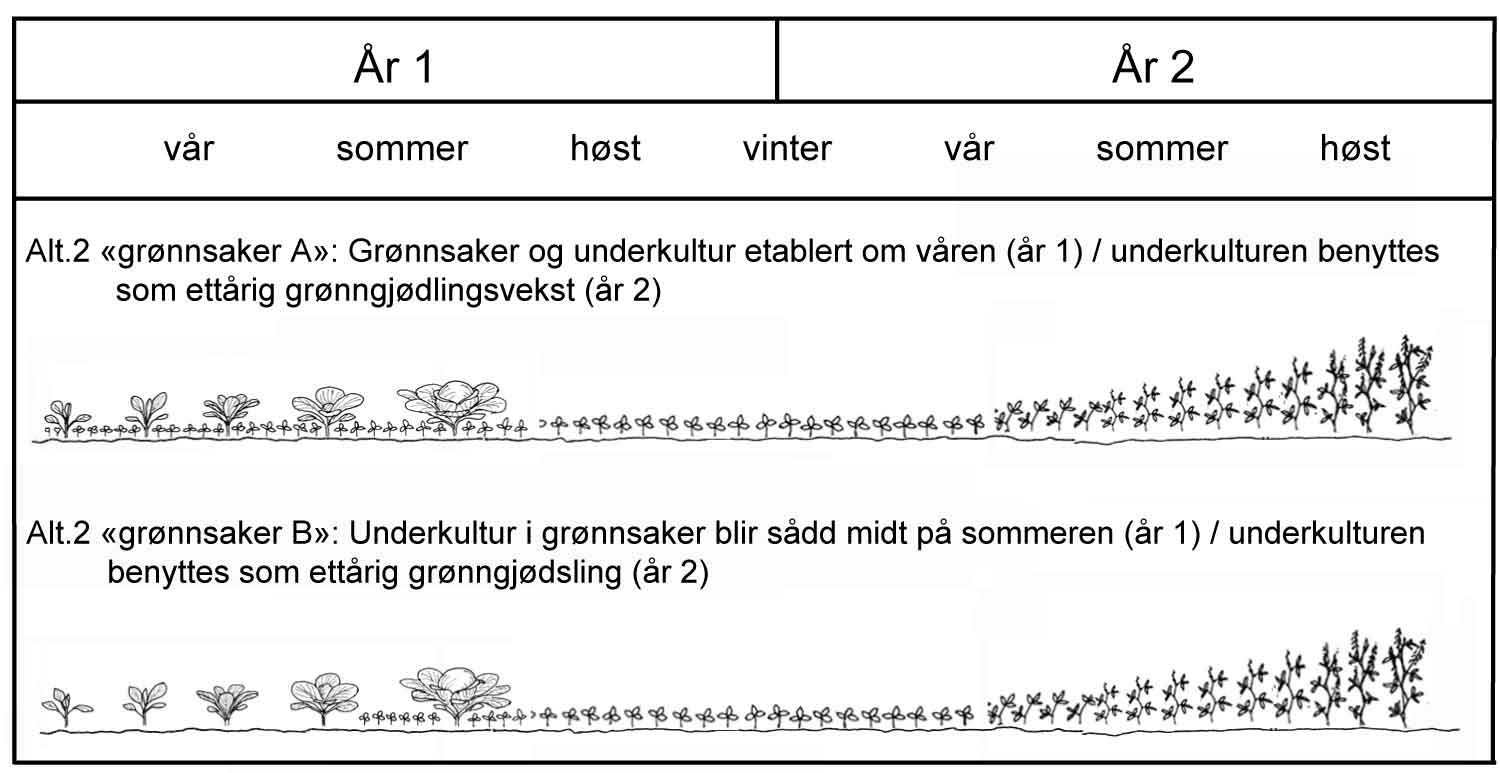
Grønngjødslingsvekster i form av ettårig grønngjødslingseng vil i prinsippet være det samme som en kortvarig eng og ha tilsvarende effekt på ugraset. Utformet på en optimal måte kan grønngjødslingsenga, i tillegg til å ha en gjødslingseffekt, være en viktig arena for sanering av flerårige ugras, for eksempel åkerdylle og åkertistel.
Etablering av grønngjødsling
Etableringen av en ettårig, eventuelt lengre grønngjødslingsperiode, kan enten starte med at man om våren sår inn den ønskede arten, eventuelt en blanding av flere arter, (se figur 3.3 alt. 1) eller at grønngjødslinga blir etablert som underkultur (gjenlegg) i korn året i forveien (se figur 3.3 alt. 2). Det er vanskelig å gi et generelt svar på hva som er best mht. ugraskontroll, svaret vil avhenge av ulike forhold på gården. Hvis man for eksempel har mye kveke vil alternativ 1 være å foretrekke fordi en da har muligheten å stubbharve om høsten. En underkultur som står igjen på jordet etter tresking, vil hindre en slik stubbharving. Fordi alternativ 2 antakelig gir best nitrogenfiksering, kan en altså bli stående med valget mellom maksimal nitrogenfiksering og ugraskontroll.
Hvis man velger å etablere en grønngjødslingsvekst om våren (alt. 1) har man også en annen interessant mulighet, nemlig å gjennomføre en brakkingsperiode om våren og forsommeren før man sår (figur 3.5). Dette er spesielt aktuelt dersom man sliter med forskjellige arter av flerårige ugras. Som vi allerede har vært innom, er alle flerårige ugras i aktiv vekst på denne tida, og man kan forvente god effekt av kombinasjonen jordarbeiding og etterfølgende innsåing av en konkurransesterk grønngjødslingsvekst. Hvis våronna starter i begynnelsen av mai, og en planlegger å så grønngjødslingsveksten ved St. Hans-tider, rekker man 3-4 jordarbeidinger med 2-3 ukers mellomrom før såing. Det vil lønne seg å avslutte brakkingsperioden med dyp pløying. De svekkede rotskuddene vil da få problemer med å komme opp til jordoverflaten.
En brakkingsperiode om våren og forsommeren vil dessuten sjelden ha store negative konsekvenser for erosjon og avrenning.

Grønngjødslingsarter og blandinger
Bonden har et sett med faktorer som avgjør valg av dyrkingssystem, grønngjødslingsvekst eller blanding, blant dem mengde nitrogen som samles, effekt på jordstruktur og ugraseffekt.
De fleste belgvekstarter er relativt trege med å danne et lukket bestand. Rett etter såing er derfor mange grønngjødslingsvekster svake i konkurransen med ugraset, og dette gjør at det ofte kan være lurt å så en blanding hvor belgveksten(e) sås sammen med mer rasktvoksende arter. Det finnes mange muligheter, men vi har i forsøk god erfaring med en blanding bestående av rødkløver (0,5 kg / dekar), italiensk raigras (1 kg), honningurt (0,5 kg) og fôrvikke (8,0 kg). Her vil honningurt og fôrvikke være de raskeste til å etablere seg, etter nedkutting vil disse gjøre mindre av seg, og rødkløveren og raigraset vil etter hvert ta over.
Hvis grønngjødslingsenga starter med underkultur i korn (alt. 2 i figur 3.3) kan man ikke så inn aggressive arter som honningurt eller fôrvikke, men det er det heller ikke behov for. Kornet selv konkurrerer ofte godt med ugraset. Man kan velge å så inn rødkløver eller hvitkløver i renbestand, eller benytte en blanding.
Ønsker man å få fangveksttilskudd, må belgvekstene ikke overskride 10 % på vektbasis. En blanding som gjør det, men som like fullt er aktuell, kan bestå av timotei (50 %), engsvingel (30 %), rødkløver (15 %) og hvitkløver (5 %).
I tabell 3.5 vil du finne opplysninger om veksthastighet, dvs. hvor raskt jorda dekkes, for enkelte aktuelle arter.
Tabell 3.5 Noen grønngjødslingsveksters evne til å dekke jorda
| Veksthastighet (utvikling) | Arter | |
| Belgvekster | Ikke-belgvekster | |
| Trege | Rundbelg1 | Rødsvingel |
| Tiriltunge1 | ||
| Jordkløver | ||
| Hvitkløver | ||
| Alsikekløver | ||
| Middels | Legesteinkløver | Rybs og raps |
| Aleksandrinerkløver | Italiensk raigras | |
| Sneglebelg | Timotei | |
| Luserne | Flerårig raigras | |
| Rødkløver | ||
| Lupin (gul, blå, hvit)1 | ||
| Perserkløver | ||
| Blodkløver | ||
| Raske | Vintervikke/lodnevikke | Reddik1 |
| Sommervikke/forvikke | Honningurt | |
| Flatbelg1 | Italiensk og westerwoldsk raigras | |
| Ært, grå foder1 | Sennep1 | |
| Havre |
1 Arter vi ikke har hatt med i egne forsøk, bestemmelse av veksthastighet bygger på erfaringer fra Danmark (Holmegaard 1987)
Behandlinger i grønngjødslingsenga
På grunn av frøugraset vil det ofte en stund etter såing være nødvendig å kutte ned grønngjødslingsveksten. Dette kan også være en fordel for belgvekstene i en blanding slik at disse etter hvert får bedre vokseplass. En art som honningurt dekker jorda raskt, men vil ha liten gjenvekstevne hvis den blir slått 1,5 til 2 måneder etter såing. Rødkløver i blandingen blir da ikke hemmet, og sikrer dermed en bedre nitrogenfiksering.
I utgangspunktet bør nedkutting gjennomføres relativt tidlig for å hindre at frøugras rekker å produsere modne frø. Et annet aspekt er at noen belgvekster etter hvert som de blir eldre, tåler nedkutting dårligere. Dette gjelder bl.a. legesteinkløver. I svensk litteratur blir det gitt en generell anbefaling om å kutte ned planten når den er ca. 30 cm høy, og ikke bruke mindre stubbehøyde enn 10 cm. Bildene i figur 3.6 viser rødkløver, legesteinkløver, luserne og en grønngjødslingsblanding bestående av rødkløver, italiensk raigras, honningurt og sommervikke, kuttet ned til ulike tidspunkt.

Under avsnittet om etablering av grønngjødslinsgvekster var vi litt inne på hvilke muligheter en har om høsten til kontroll av flerårige ugras hvis grønngjødslingsenga etableres som underkultur (alt. 2 i figur 3.3). Den muligheten en har for å kontrollere kveka, i alle fall til en viss grad, om høsten i en underkultur, er å kutte kveka (og grønngjødslings-veksten) når den senest er på 3-4-bladstadiet. Hvor god effekt dette har på kveka, og hvor mye det går utover nitrogenfiksering og overvintringsevne, vet vi ikke sikkert i dag. Med mye flerårig ugras i underkulturen om høsten er det nok lurt å slå ned disse en eller flere ganger. Spesielt gjelder dette hvis man har tresket tidlig og i tillegg får en lang og fin høst etterpå. Figur 3.26 viser hvilket potensiale kveka har til å bygge seg opp utover høsten hvis den får stå uforstyrret.

Et annet aspekt om høsten er hvordan nedklippingen bør gjøres for å sikre en best mulig overvintring for grønngjødslingsvekstene. Det er vanskelig å gi et godt svar på dette, men et generelt råd er at en bør slå så tidlig som mulig. I Sør-Norge helst før midten av september, slik at plantene kan ha noe tid til å akkumulere karbohydrater til vinteren. Kulturplantene bør generelt forstyrres minst mulig om høsten.
Siden mange som driver økologisk, sliter med kontroll av åkertistelen, skal denne arten vies litt ekstra oppmerksomhet. Det er interessant å merke seg at mange av de anbefalingene som vi gir i dag for kontroll av dette ugraset, faktisk ligner mye på de rådene som ble gitt for svært lenge siden. Allerede i 1848 utarbeidet amerikaneren Stevens følgende liste med gode råd i kampen mot åkertistelen:
Vi ser altså at eng til slått gjennom mange år har vært ansett å være effektiv i åkertistelkampen. Hvilken effekt slåtten gir, er avhengig av slåttetidspunkt, antall slåtter og engas botaniske sammensetning.
Forsøk har vist at rødkløver og luserne kan konkurrere sterkt med åkertistel. Som vi har vært inne på tidligere, vil optimalt tidspunkt for nedkutting være når åkertistelen er svakest, dvs. på 8-10-bladstadiet. Kontroll av åkertistel blir betydelig lettere om et omløp inneholder eng over flere år. Forsøk i Amerika har vist at enga må ligge i tre år for at effekten på åkertistel skal være god. Antall engår er imidlertid ikke den eneste avgjørende faktoren, totalt antall år i omløpet og hvilke andre vekster omløpet inneholder, er også viktig. En svensk tommelfingerregel sier at det blir problemer med åkertistelen hvis engandelen kommer under 40 %, for eksempel mindre enn 2 av 5 år. Spesielt gjelder dette hvis de andre årene av omløpet er dominert av korn. Behovet for eng for å sanere flerårige ugras blir redusert hvis man har andre kulturer i omløpet, hvor det også går an å drive kontroll, for eksempel radrensing.
Anbefalinger sier at enga bør slåes minst to ganger i vekstsesongen, men gjerne tre ganger, for å oppnå god effekt mot åkertistel. Forsøk i Sverige der rødkløvereng ble slått med intervaller på 1, 2, 3, 4, 5, og 6 uker viste at slått hver fjerde uke gav best effekt (se figur 3.8). En årsak til at 4 ukers intervall gav best resultat, var at rødkløveren da tålte slåtten uten at det gikk ut over gjenveksten. Senere forsøk har dessuten vist at det er de første slåttene om sommeren som hemmer åkertistelen mest.
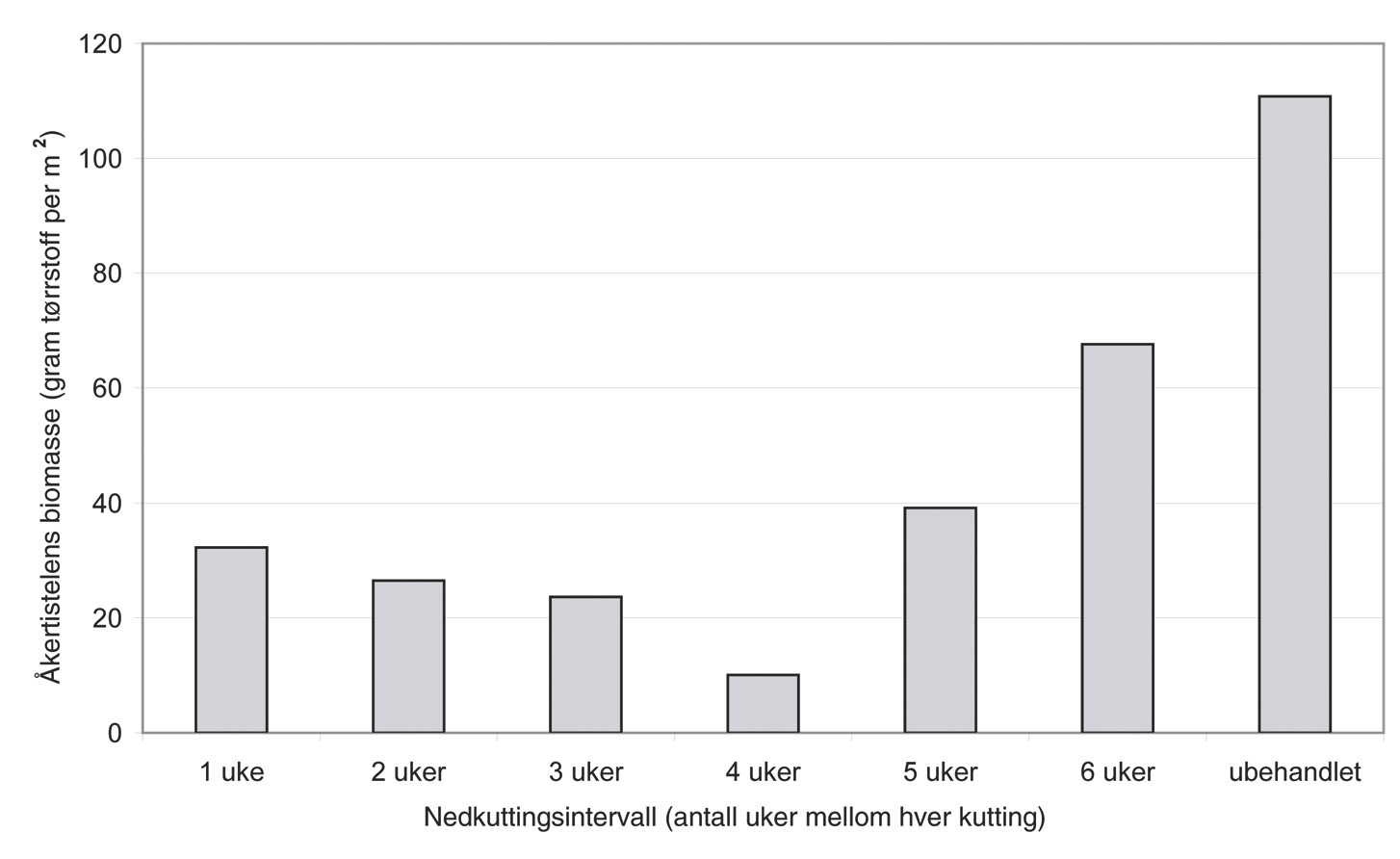
Sammenlignet med åkertistel finnes langt mindre kunnskap om hvilke effekter eng har på åkerdylle. Imidlertid vil mye av det som er skrevet om åkertistel, også gjelde for åkerdylle. En god bekjempelsesstrategi vil også her bestå i at man kutter ned åkerdylla når tørrstoffmengda er minst, dvs. på 5-7-bladstadiet. Antakelig vil åkerdylla utarmes raskere i enga enn hva tilfelle er for åkertistel. Undersøkelser i Norge viser da også at åkerdylla er mindre vanlig i eng enn hva tilfelle er for åkertistel (se tabell 3.4).
En ulempe med nedkutting av et grønngjødslingsbestand er at nitrogen i plantematerialet på jordoverflaten tapes som ammoniakkgass til lufta. En mulighet for å nyttiggjøre seg dette plantematerialet (og nitrogenet) kan, som nevnt tidligere, være å bruke det som dekke i grønnsaker på et annet skifte.
Undersøkelser i Sverige har vist at forekomsten av kveke er ganske lik for de forskjellige kornartene, uavhengig av om det var vår- eller høstkorn. Det er også svært interessant å merke seg at det ikke ble funnet store forskjeller om kornartene ble dyrket i omløp med eng eller i omløp uten eng. Det er imidlertid i undersøkelser funnet at mengde kveke minsker med alder på enga når denne ligger i mange år. Brakking og jordarbeiding har vist større virkning på kvekebestanden enn drifta av enga. Forsøk med raigras i Norge har imidlertid vist at denne arten kan konkurrere bra med kveka.
Figur 3.3 viser to ulike måter å etablere en «mellomkultur»:
«Alt. 3»: Så for eksempel hvitkløver eller rødkløver som en underkultur i korn. Underkulturen pløyes inn i jorda enten senhøstes eller våren etterpå. Det kan være en mulighet å slå ned underkulturen om høsten for å behandle ugraset som vokser i denne, men vi vet ikke hvor god effekt på ugras dette gir.
«Alt. 4»: Så inn en «mellomkultur» om høsten etter at kornet eller en annen tidlig kultur (tidligpotet, grønnsaker) er høstet. En flerårige belgvekst som rødkløver kunne være aktuell, men som regel vokser den for tregt til å passe for denne bruksmåten. Det har blitt gjennomført ganske omfattende undersøkelser i Norge for å finne velegnede arter og sorter til bruk som «mellomvekst» (tekstboks 3.4).
Dersom hovedkulturen blir høstet tidlig nok, er det aktuelt å så inn en belgvekst, eventuelt en blanding, om høsten. Et grønt dekke gjennom høst, vinter og vår kan ha flere fordeler, bl.a. samle nitrogen til etterfølgende kultur og konkurrere med ugraset. Under følger en liten gjennomgang av erfaringer som er gjort med noen vinterettårige og toårige belgvekstarter.
Jordkløver (Trifolium subterraneum)
Jordkløver må ikke såes før midtsommer om den skal kunne overvintre. Etter overvintring blomstrer den allerede i mai, og naturlig nedvisning starter tidlig på våren/forsommeren. Vår idé med denne arten har vært at det utover våren og forsommeren skulle være mulig å plante grønnsaker direkte (eventuelt lage et plantebed ved hjelp av seksjonsfres) i det døende jordkløverbestandet. Omfattende undersøkelser av overvintringsevne for å finne aktuelle jordkløversorter for Norge har gitt nedslående resultat. Best overvintringsevne viste 'Denmark', fulgt av 'Nuba' og 'Mount Barker'. Livssyklus og voksemåte peker ut jordkløver som nær sagt den «ideelle» underkultur for bruk i radkulturer, men den dårlige overvintringsevna har gjort at det ikke arbeides mer med denne arten. Spesielt interesserte bønder eller småhageeiere langt sør i landet kan jo gjøre et forsøk.
Lodnevikke/vintervikke (Vicia villosa)
Lodnevikke er en langt mer velegnet art for norske forhold enn jordkløver, men bør antakelig bare dyrkes i Sør-Norge, men heller ikke der i innlandet. Sorten 'Hungvillosa' har vært best, men 'Welta' og 'Villana' har fulgt 'hakk i hæl'. Lodnevikke bør ikke såes for tidlig på ettersommeren, siste halvdel av august eller begynnelsen av september synes å være beste såtid. Fordi man bør bruke 7-8 kg såfrø per dekar for å få god dekning, blir høye frøutgifter et minus for lodnevikke. Ved vellykket etablering danner lodnevikke et svært tett bestand som konkurrerer godt med ugraset. Lodnevikke har god tilvekst om våren og forsommeren, og samler dermed mye nitrogen på denne årstida. Vi anbefaler derfor spesielt å bruke lodnevikke som forkultur til litt senere hold av grønnsaker. Nitrogen som finnes i biomassen til lodnevikke frigjøres raskt etter nedmolding.
Blodkløver (Trifolium incarnatum)
Av sortene vi har testet har 'Heusers Ostsaat' vist suverent best overvintringsevne. Vi skal ikke komme så detaljert inn på denne arten, mye av det som står om lodnevikke kan overføres til blodkløver. Sammenlignet med lodnevikke har imidlertid blodkløver visse svakheter: Litt dårligere overvintringsevne, mer glissen vekst og dermed ikke så god konkurranseevne mot ugraset, og dessuten, i et næringsforsyningsperspektiv, den brytes saktere ned. Et par fordeler skal imidlertid også nevnes: De blodrøde blomstene er et usedvanlig vakkert syn for øyet, og frøkostnadene er langt lavere. Men alt i alt er nok lodnevikke å foretrekke framfor blodkløver for de fleste bruksområder.
Legesteinkløver/gul steinkløver (Melilotus officinalis)
Våre forsøk mer legesteinkløver har bare omfattet en sort, 'Nordgold Yellow', og mye tyder på at det ikke er noe stort behov for andre og bedre sorter. Til og med «ekstremvinteren» 2000/2001 på Østlandet, overlevde denne arten godt. Den gode overvintringsevna er kanskje ikke så overraskende, denne arten finner vi nemlig viltvoksende langt nordover i landet. I motsetning til de andre artene vi har omtalt, er legesteinkløver en toårig art. Våre forsøk har vist at legesteinkløver trives best ved såing om våren og litt utover forsommeren. Overvintringsevna blir betydelig redusert ved etablering på ettersommer eller høst. Ved tidlig såing vil legesteinkløver blomstre første høsten, men dette betyr ikke at den går ut. Andre våren dekker legesteinkløver jorda raskt. Får den stå urørt utover forsommeren vil den bli svært høy, gjerne 2 m. Et viktig aspekt ved bruk av denne arten er at hvis den ikke tidligere har vært dyrket på åkeren, må såfrøet smittes med en bakteriekultur. I litteraturen finnes det mange utsagn om at legesteinkløver med sitt dype rotsystem har svært god effekt på jordstrukturen.

Bruk av underkultur i ulike kulturvekster har en ugraseffekt. Dette er vist både i grønnsaker (ikke mye brukt på grunn av skadelig konkurranse med kulturen) og i korn. De fleste undersøkelsene har konkludert med at effekten av underkultur er best mot frøugras, men virker også litt på flerårige arter. En undersøkelse viste for eksempel at underkultur i havre i gjennomsnitt halverte kvekemengda i forhold til havre uten underkultur. På den annen side kan situasjonen fort bli motsatt hvis ugraset, i alle fall kveke, allerede er etablert i åkeren. Underkulturer og fangvekster vil i stor grad vanskeliggjøre mekaniske tiltak som stubbarbeiding om høsten.
Bruk av underkulturer eller fangvekster før hovedkulturene er sådd, og etter at de er høstet, kan være et viktig bidrag til å hindre at ugras etablerer og formerer seg på tider av året da jorda skulle ligge «svart». Hvilke vekster som bør brukes, og eventuell behandling av disse, har vi i dag for lite kunnskap til å si noe generelt om.
Jordarbeiding mellom kulturvekstene i et omløp har flere funksjoner, som tillaging av så- eller plantebed, innarbeiding av gjødsel og plantemateriale i jorda, og kontroll av ugras. For alle driftsformer er det dessuten viktig at jordarbeidinga utføres på en slik måte at man oppnår best mulig jordstruktur, samt minst mulig erosjon og utvasking. Å unngå jordpakking er spesielt viktig. God jordstruktur vil i seg selv være gunstig i ugraskampen fordi ugraset ofte greier seg bedre enn kulturplantene på strukturskadet jord.
Det finnes mange gode argumenter for redusert jordarbeiding, men så godt som all erfaring i dag tilsier dessverre at det er et motsetningsforhold mellom redusert jordarbeiding og ønske om mindre herbicidbruk. Spesielt gjelder dette flerårige ugras som veldig lett tar overhånd ved redusert jordarbeiding. Svært mange undersøkelser viser hvor viktig god jordarbeiding er, inklusivt pløying, hvis ugraset skal kontrolleres uten bruk av herbicider.
Jordarbeiding er viktigere i økologisk drift enn i konvensjonell drift. Jordarbeidinga skal i stor grad kompensere for bortfallet av kjemiske ugrasmidler, men den har også stor virkning på frigjøring av plantenæringsstoffer. Dette har i neste omgang betydning for veksten av både kulturplanter og ugras. Skjer jordarbeidinga på feil tidspunkt, kan den føre til tap av næringsstoffer. Jordarbeiding og kjøring på jorda er med på å ødelegge jordstrukturen, mest når det er rått. Derfor skal en ikke kjøre mer en det som er nødvendig, og passe på å unngå kjøring når det er rått. Sagt på en annen måte: En skal bruke minst mulig energi til å ødelegge jorda, og mest mulig til å få et godt resultat. Dette er, som nemnt, viktigere i økologisk drift enn i konvensjonell drift. I korndrift, hvor en har mye åpen åker, blir det mer jordarbeiding enn der eng dominerer.
Dersom jorda er pakket, blir det dårlige forhold for kulturplantene. Dette gjør at de blir dårligere til å konkurrere med ugraset. En del ugras trives bedre enn kulturplantene i pakket jord. Alle har vel sett hvordan tunrapp har bredt seg i åkrene, særlig på vendeteiger og langs kantene, fordi jorda har blitt mer og mer pakket. Ugras som har dypt og kraftig rotsystem, vil også konkurrere meget godt. Disse kan utnytte dypere jordlag som ikke er pakket, i motsetning til kulturplantene med grunnere rotsystem.
Belgvekstene som er avgjørende for å få økologisk drift til å fungere, lider ved jordpakking og dårlig jordstruktur. Disse plantene er avhengig av Rhizobiumbakterien og denne er igjen avhengig av luftveksling for å få både oksygen og nitrogen.
Hvordan unngå jordpakking og ødelegging av jordstrukturen:
Dekk og lufttrykk i dekkene
Pakketrykket like under et hjul (i de øverste 15-20 cm) er direkte avhengig av lufttrykket i dekket. For å få lite pakketrykk, må lufttrykket i dekket være lavt. Det er videre en sammenheng mellom maskinvekt, dekkdimensjon og lufttrykk. Jo tyngre en maskin er, jo høyere må lufttrykket i dekkene være, dersom ikke dekkdimensjonen økes. Både diameter og bredde har betydning. En økning i diameteren betyr likevel relativt lite. Dessuten er det begrenset hvor mye diameteren på hjulene på en maskin kan økes uten at det skaper problemer. Det er enklere å øke bredden. Setter en på dekk som er 30-40 % bredere, kan en senke lufttrykket til omtrent det halve. (En må se etter i trykk- og belastningstabeller hvilke lufttrykk en bør bruke, fordi det er variasjon på dekktyper, spesielt når en går over til lavere profil. Det er også en grense nedad hvor lite trykk et dekk kan tåle.)
Maskintyngde
Jo tyngre maskin, jo større lufttrykk må en bruke, dersom en ikke går opp i en dekkdimensjon som kompenserer for tyngden. Under en lett maskin reduseres trykket raskt nedover i jordlagene. Under en tung maskin, dvs. en maskin med aksellast over 4-6 tonn, holder trykket seg nesten konstant i de øvre 15-20 cm (jo råere, jo lengre ned). Fordi jorda nedover som oftest er fuktig, spesielt i våronna, vil et lufttrykk i dekket på over 0,6-0,8 bar (1 bar = 14 pund per kvadrattomme) være grensen for når det oppstår skadelig pakking i undergrunnen. Spesielt vil dette kunne skje når en pløyer og kjører i fåra. Pakking under plogsjiktet gir relativt små årlige avlingsutslag, men det kan ta mange tiår før jorda løses opp igjen. Skal en være sikker på at en ikke får skadelig jordpakking, burde en i økologisk dyrking ha aksellast under 4000 kg og dekk som kan gå med 0,6 bar.
Jordfuktighet
Jo større jordfuktighet, jo lettere lar jorda seg pakke. I økologisk drift bør en derfor være meget påpasselig med å unngå å kjøre når det er rått.
Spor
I forbindelse med all jordarbeiding og annen kjøring på jordet bør hjulutstyret være så stort at en unngår spor som skaper problemer i etterfølgende arbeid.
Hensikten med pløying er å:
Tradisjonelt har en pløyd om høsten. Imidlertid har bevisstheten om erosjon og næringstap gjort at det blir stilt spørsmål ved om dette er riktig. Høstpløyinga har derfor gått sterkt tilbake. Økonomisk kompensasjon for ulemper med andre former for jordarbeiding, og tilskudd til fangvekster har bidratt til endringen. Der hvor det normalt er stabil vinter med tele, kan relativt sen høstpløying være gunstig fordi en setter i gang nedbryting og frigjøring av næringsstoffer, mens utvaskingen blir relativt beskjeden. Det er viktig å vente med vårpløyinga til det har tørket opp for å unngå jordpakking.
Bruksanvisningen til plogen gir en rekke tips om innstilling, noen av de viktigste er:
En tyngre plog holder jevnere pløyedybde. Derfor er en vendeplog bedre enn en teigplog. Med vendeplog slipper en dessuten rygger og aurfårer, noe som er gunstig med tanke på ugrasharving og radrensing.
Veltefjøl
Ofte kleber jord seg fast til veltefjøla og ligger i et 10-20 cm tykt lag. Plogen mister da evnen til å snu velta og dekker dermed ikke ugras og planterester. Resultatet blir dårlig såbed, mye rotugras og liten avling. Heldigvis er det flere muligheter for å pløye bedre på slik jord. Noen av disse løsningene koster mer enn en vanlig veltefjøl av stål, men det blir som oftest småpenger sett i forhold til avlingstapet. Dette problemet blir derfor mer omfattende behandla fordi det er lite publisert på dette området. Litt forenklet kan vi si at det i jorda finnes partikler som virker som små magneter. Disse trekkes mot stålet på veltefjøla. Det samme gjelder også vannmolekylene. Når en har en viss fuktighet, avhengig av jordtype, suger jorda seg fast til veltefjøla. Er jorda fuktig, vil vann presses ut av jorda i overgangen mellom jorda og veltefjøla og danne en vannfilm. En ser det på pløgsla, velta blir blank. Det dannes flere lag med vannmolekyler som virker som et lag med kuler slik at jorda glir lett langs fjøla. Spesielt på myrjord og jord med mye organisk materiale blir ikke trykket mot fjøla stort nok til at det presses ut vann. Jorda er for lett. Derfor er slik jord problematisk. Skal det på denne jordtypen dannes vannfilm, må jorda være meget rå. Er på den annen side jorda tilstrekkelig tørr, vil jordpartiklene heller ikke greie å suge seg fast på veltefjøla. Dette kan en se når en pløyer tørr myr. Når vanninnholdet ligger mellom disse to ytterpunktene, kleber jorda seg lett til veltefjøla. Med andre ord, en kan oppleve å få fin pløgsle på problemjord i år hvor det er meget tørt eller meget rått, og en uryddig pløgsle ellers.
Fordi det er vanninnholdet og trykket mot veltefjøla som bestemmer om det skal bli vannfilm, kan det i noen tilfeller gå bedre dersom en øker hastigheten over problemflekker. Når hastigheten øker, øker trykket mot fjøla. Kjører en med lang toppstang og plogåsene heller mot velta, vil bakre enden av veltefjøla presses ned i velta, velta stukes, trykket lengre fram på veltefjøla blir for lite og jorda glir ikke. Av og til hjelper det da å korte inn toppstanga litt og korte inn avvatringsstaget på en teigplog, og justere stoppeskruen(e) på en vendeplog.
Når veltefjøla ruster, dannes det først rust i punkter der stålet har mikroskopiske porer. Disse porene blir etter hvert større og selv om ei veltefjøl med gravrust virker tilsynelatende blank, er det små ujevnheter som gjør at jorda suger seg bedre fast. Har en problemjord, er første bud at en ikke skal la plogen stå og ruste. Sett den derfor under tak og/eller ha på rustbeskytter (samme stoff som brukes i kanaler på biler). Husk at olje og diesel er lettere enn vann og renner av i regnvær. Gammel motorolje inneholder syrer som kan øke rustingen. Har en fått rust i fjøla, hjelper det å bruke skiver med smergellerret på vinkelsliperen eller kjøre den ren på slipende jord, men fullgodt blir det aldri. Skiva med smergellerret er også fin å bruke til å polere et nytt skjær. Da unngår en at lakk og ujamheter på skjæret blir årsak til at jorda kliner seg på.
De fleste ploger leveres i utgangspunktet med vendbar spiss som ligger oppå skjæret. Dette er en god løsning sett fra et slitasjesynspunkt, men kan være en medvirkende årsak til at det kleber på fjøla. Bak denne spissen mister velta kontakt med skjæret. Presset mot stålet blir for lite, det blir ingen vannfilm og det legger seg jord bak spissen. Dette er som oftest utgangspunktet for klebing på fjøla. Her finnes det to løsninger som er bedre. Alle plogfabrikantene kan levere skjær hvor skjær og spiss utgjør en enhet. Noen plogfabrikanter, blant andre Kverneland, kan også levere skjær med skiftbar, ikke vendbar spiss. Overflaten på denne spissen ligger i samme plan som skjæret, og går derfor bedre i klebrig jord enn når spissen ligger oppå. Slitasjemessig er det en god løsning fordi spissen kan skiftes.
Etter vår mening burde en alltid bruke forplog eller skumskjær når en pløyer. Skumfjøla har blitt populær, men skummer dårligere. Dersom en har jord som kleber på veltefjøla vil skumfjøla forsterke problemet fordi den løfter velta i forhold til fjøla. Presset mot veltefjøla blir da mindre slik at det ikke dannes vannfilm.
Plastveltefjøl
Har en plastveltefjøl og skjær uten ovenpåliggende spiss, vil en neppe få problemer med klebing på fjøla siden vann og jord ikke suger seg fast til plasten.

Plast er derimot ikke så slitesterkt som stål på steinholdig jord. En får derfor større utgifter fordi fjøla slites ut raskere og koster mer enn stålfjøl. Her er det viktig at en ikke ser seg blind på denne kostnaden og glemmer at en vil få mindre rotugras og større avling. Kveka kan en nok bekjempe med høstbrakking, men det koster også penger i form av arbeid, traktorbruk, slitasje av harveutstyret og ikke minst tapt nitrogen og ekstra jordpakking. På Blæstad (Høgskolen i Hedmark) har en prøvd plastfjøl på jorder som skifter mellom ekstremt slitende jord og klebende myrjord. Plastfjøla har like god form som stålfjøler og gir derfor topp pløyeresultat i all jord. Erfaringene på denne jorda kan tyde på at plastfjøla holder omtrent like lenge som to skjærsett (og fire vendbare spisser). Trolig kan vi med en treskjærs vendeplog pløye ca. 1000 dekar på slik jord. Seks veltefjøler koster ca. 10 000 kroner. Dette betyr at slitasjen på veltefjøler koster ca. kr 10 per dekar. Dette tilsvarer kun 3-4 kg økologisk korn eller 2-3 kg økologisk kraftfôr. De som har Kverneland plog trenger ikke å skifte hele plogen, det holder med å kjøpe nye kropper.
Stripekropp
Alle plogfabrikkene leverer i dag stripekropper. Her er hele stålfjøla erstattet med tykke stålspiler som er 5-10 cm brede og med mellomrom som er omtrent like store.

Det er to årsaker til at stripekroppen går bedre på klebrig jord. Det blir større trykk på de smale stripene, slik at det lettere oppstår vannfilm og jorda glir lettere. Selv om det kleber seg jord på spilene, blir det ikke så tykt lag som på ei hel veltefjøl. Stripekroppen vil derfor tilnærmet beholde sine velteegenskaper under slike forhold. Stripekroppen er i utgangspunktet konstruert for å selges til områder i andre land hvor en kun vårpløyer åker. Den er brattere enn de vanlige plogene, og dette medfører at velteegenskapene i vollpløgsle ikke er riktig så god som hos vanlig stålfjøl. Velteegenskapene er imidlertid enormt mye bedre enn ei stålfjøl med et tykt lag jord. Stripekroppen er derfor utmerket på åker, og et kompromiss på vollpløgsle på jord med mye stein. Vi har ikke erfaring med hvor fort stripene slites ut, men antakelig blir kostnadene omtrent de samme som med en vanlig stålfjøl. I innkjøp koster imidlertid stripekroppen litt mer. På Kverneland ploger kan en kjøpe stripekropp til eldre ploger.
Utstyr på plogen som har betydning for ugrasregulering
Dagens ploger kan leveres med skiveristel eller knivristel, forplog eller skumfjøl. Alle disse er mer eller mindre egnet til ugrasbekjempelse. Det er uansett viktig at plogen har slikt utstyr.
Ristelen skal skjære av horisontale røtter og sprette opp fåra for plogen (det siste gjør at plogen går stødigere). Skiveristelen (rulleskjær) er best når det gjelder å skjære av røttene. Knivristelen (skjærkniv) vil rive av røttene slik at for eksempel jordstenglene hos kveke ofte blir stikkende ut i kanten av velta. Dersom en delvis høstpløyer, bør en velge skiveristel. Har en seig, fuktig halm vil en glatt skiveristel lett stoppe, og halmen skyves foran skiveristelen. Under slike forhold skal en bruke takkete skiveristler. «Tannhjulseffekten» gjør at takkete skiveristler går rundt.
Når velta snus rundt vil lufta presses foran velta. Dette fører til at uten skummeutstyr blir grønne plantedeler av ugras og stubb fra foregående vekst bøyd opp mellom veltene. Ugraset kan da vokse videre og sjukdomsmitte overføres direkte til neste års kultur, dersom den er mottakelig for samme sjukdom. Sjukdomsoverføring er ikke noe stort problem dersom en driver effektivt vekstskifte. Skummeutstyret skal fjerne grønne plantedeler og planterester på toppen av velta. Blant alternativene som finnes, er forplogen klart å foretrekke. Forplogen (skumskjæret) skjærer av en trekant på den øvre del av velta (figur 3.13). Planterester etter foregående avling kastes ned i fåra og dekkes.
Det er viktig å ha riktig dybde på forplogen. Går den for grunt, vil det fortsatt stikke opp grønne deler, og går den for dypt, vil det bli vanskelig å tette mellom veltene, spesielt på stiv jord. Får en ikke tettet mellom veltene ved høstpløying, får en lys ned til ugraset, og det fortsetter å vokse. Det er meget viktig at forplogen er blank og glatt når jorda har en tendens til å klebe på. Før bruk må en fjerne lakken og pusse overflata blank med slipepapir på vinkelsliperen.
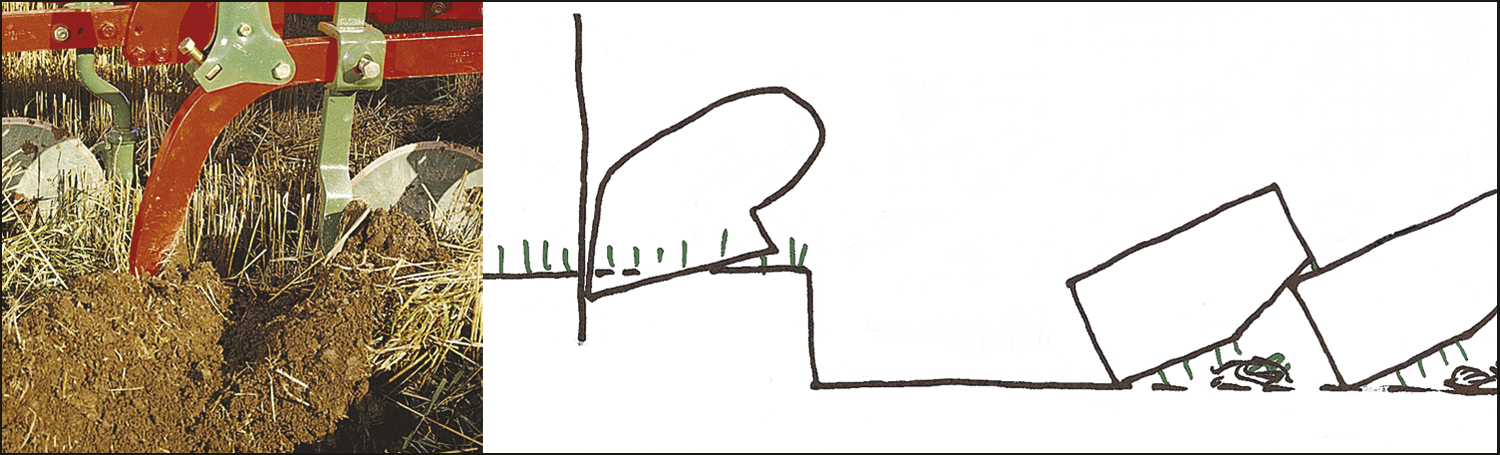
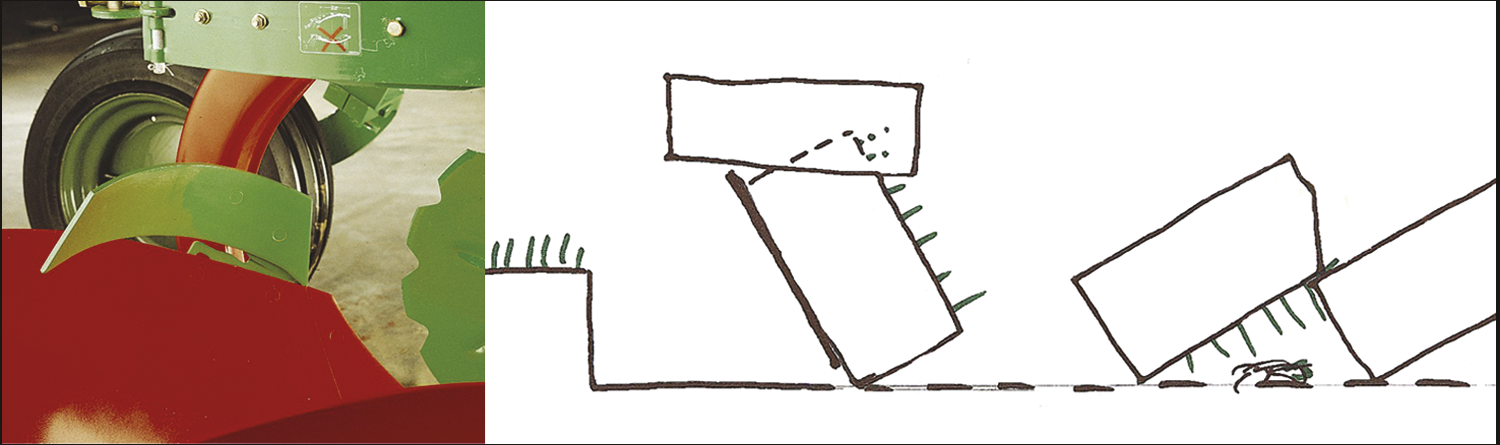
Skumfjøla (skumvingen) skraper av en annen trekant («feil» trekant) av velta, se figur 3.12 og figur 3.13. I lett jord og åker vil toppen av velta brekke av og legge seg ned mellom veltene, dekke grønne plantedeler og tette mellom veltene. På stivere jord skjer ikke dette. Der vil grønne plantedeler og halmstubb fortsatt stikke opp mellom veltene. På voll og dels på grønngjødslingsarealer vil rotsystemet armere jorda nær overflata. Da vil ikke skumfjøla klare å skrape vekk de grønne plantedelene. En kan derfor konkludere med at i økologisk drift bør en velge forplog. Er det mye halm som i tillegg er dårlig kuttet, vil skumfjøla gå noe bedre enn forplogen. For å få best mulig ugraskamp med plogen bør en sørge for at halmkutteren kutter godt (slipe knivene) og sprer halmen godt. I økologisk drift burde ikke halmen være noe problem, men går ikke forplogen, er skumfjøla bedre enn ingenting.
Hvordan pløyinga legges opp i praksis
En skal alltid kjøre markeringsfår for vendeteigene slik at en setter i og løfter opp på ei linje som er parallelt med åkerkanten. Dersom en kjører ulikt ut mot vendeteigen, blir det enten igjen upløyd areal eller det blir dobbeltpløyd når en pløyer vendeteigen. Helst bør en måle ut avstanden til åkerkanten slik at bredda på vendeteigen går opp med bredda på plogen. Får en igjen ei halv plogbredde vil det bli dobbeltpløying og dårlig ugraskamp. Rotugras i kanten av jordet er ofte «smittekilden» for resten av åkeren.
Når en begynner ved kanten av jordet, enten en pløyer vendeteig eller begynner med vendeplog, bør en først kjøre ei får med bakre plogen til halv dybde og velte vekk fra kanten, se figur 3.14. Denne velta skal igjen veltes inn mot kanten og dekkes med ei velte med første plogkroppen, se figur 3.15. Denne velta skal kjøres til ca. halv dybde, og så skal de neste to veltene ha øket dybde ned til omtrent normal pløyedybde.

En skal alltid veksle med velteretning. Velter en bare en retning, vil jorda forflyttes mot en kant. Dersom en har teigplog, skal en bestandig legge ryggen der en hadde avslutningen (aurfåra) sist en pløyde, og få avslutningen der en hadde ryggen. Dette krever at en måler opp og noterer hvor rygger og avslutning lå. I figur 3.16 er det et forslag til hvordan dette kan gjøres. Først måler en ut vendeteigene 10-15 m brede, avhengig av traktorstørrelse, rundt hele stykket. Spar ikke på vendeteigbredde. Deretter måler en opp bredden mellom vendeteigene og setter opp stikker for pløying av ryggene 1/6 av bredda inn fra hver side. Første året pløyer en to rygger, og når en har pløyd disse ryggene og teigene ferdig ut til vendeteigen, har en igjen 2/6 midt på stykket. Dette slås utover og en får igjen en aurfår midt på stykket. Vendeteigene pløyes innover dette året. Andre året lages det like brede vendeteiger og det legges en rygg midt på stykket, der aurfåra var. Så pløyer en et felt tilsvarende 1/6 ut fra hver side av ryggen. Nå står det igjen to felt på hver side som tilsvarer 2/6. Disse veltes utover mot markeringsfåra og mot ryggen. Er en nøyaktig kommer aurfåra der ryggen var sist år. Vendeteigene pløyes utover dette året. Med et slikt opplegg får en mindre problem med rygger og aurfårer, og en får mindre ugras.
Slodden skal knuse klump, planere og forberede såbedet. Har en for mye klump i det øvre sjiktet, er det vanskeligere å få løs jord til å dekke ugraset når en senere skal ugrasharve. Planeringa er viktig både for såbedet og for å få godt resultat av ugrasharvinga. Selv om dagens ugrasharver har fjærende tinder, vil de ikke gjøre god nok jobb dersom overflata er ujevn. Tindene skal kun gå 2-3 cm ned i jorda. Med tanke på radrensing, er det også viktig med jevn overflate. Uansett hvilket planeringsutstyr en bruker, er det en fordel å ha tvillinghjul, eventuelt spesielt brede dekk på traktoren, slik at det ikke blir dype spor. Slodden gjør også en forberedelse til såbedet ved at det blir jevnt i overflaten, og jevnere hardhet i jorda. Dersom en slodder like etter jorda har blitt tørr på overflaten og venter 4-6 dager med harvinga, vil det spire mye frøugras som drepes når en harver (falskt såbed).
Typer av planeringsredskaper
Ut fra et ugrassynspunkt, spiller det antakelig ikke så stor rolle hvilket planeringsutstyr en bruker, bare en får et plant jorde med henblikk på ugrasharving og radrensing.
Slodden er et rimelig og effektivt redskap. En skiller mellom understrømningsplanke og overstrømningsplanke. Understrømningsplanken er som oftest laget av kanalstål og kan stå vinkelrett på åkeren eller noe vinklet forover. Overstrømningsplanken er et vinkeljern som høvler av og løser opp fast jord. En slodd som skal planere godt, må være minst 2,5 m bred, ha tre sloddeplanker og eventuelt ha hjul bak for å få større avstand mellom trepunktsfestet og bakre bærepunkt. Det er en fordel at første sloddeplanken er en understrømningsplanke. Denne har større evne til å knuse klump mot ei urørt, hard høstpløgsle. Er det en overstrømningsplanke, et liggende vinkelstål, vil den skjære løs klumpen som dermed blir vanskeligere å knuse. Den andre sloddeplanken kan med fordel være en overstrømningsplanke, mens den tredje bør være en understrømningsplanke. Slodden må også være så tung og plankene så høye at de hele tida har nok jord med seg til å fylle (alle) hull. På jord hvor det ikke er for mye stein, kan det med fordel være pigger eller flatstål under slodden. Disse kan under gunstige forhold lage et tilstrekkelig såbed, men normalt er det en fordel å kjøre over en gang med harva. De fleste harver kan utstyres med sloddeplanke foran. Har en lett jord og er meget nøye med innstillingen av plogen, slik at den lager jevne velter, kan en slik sloddeplanke være tilstrekkelig dersom harva er tung. På tyngre jord blir likevel slik enkel slodding som oftest for dårlig. Overflata blir ikke jevn nok for ugrasharvinga. På steinholdig jord vil harva lett bli løftet når sloddeplanken møter stein. Da blir planeringa dårlig og harvedybden ujevn. Sloddeplanke er et gunstig supplement til slodden og til finplanering (etter steinfjerning).
Kombinert planerings-, pakkings- og klumpknusingsredskap finnes det i dag mye av. Disse veier mye og krever en stor, tung traktor. Selv om en her i en viss grad kan klare seg med en kjøring, blir pakkinga alt for stor der hjulene går. Lettere enkeltutstyr kjørt med lett traktor med tvillinghjul er bedre. Flere fabrikker tilbyr nå tromler med ulike former for planeringsredskap foran. Hensikten er å få planert overflata og pakket jorda passelig mye sammen etter vårpløyinga. Igjen kreves det meget jevn pløying dersom en ikke slodder på forhånd.
Kjøremønster ved slodding
Det er viktig at en tenker på ugrasspredning når en slodder og harver. En bør for eksempel unngå å kjøre slik at slodden for hver tur er bortom ytterkanten av vendeteigen. I ytterkanten kryper nemlig rotugraset ut over jordet. Slodden og harva kan da dra formeringsorganene utover jordet. Slodd derfor først langs kanten rundt hele stykket, slik at vendeteigen og overgangen til jordet er sloddet. Slodd så selve stykket. Er slodden god, kan en kjøre på langs av pløgsla, spesielt på nypløyd åker. På stivere jord og høstpløyd åker, kan det lønne seg å kjøre litt diagonalt. Vendeteigene sloddes til slutt på nytt.
Det finnes et utall av redskaper for jordarbeiding, men vi konsentrerer oss om harver som er av direkte eller indirekte interesse for ugrasregulering i økologisk jordbruk. Også når det gjelder jordarbeidingsredskaper kan en snakke om «moter». Ikke alt som er nytt er automatisk det best egnede for økologisk drift. I den senere tid har det kommet mange kombinasjonsredskaper som er effektive, men som trenger en meget stor traktor. Selv om en reduserer antall kjøringer, vil dette kunne medføre så stor jordpakking at de er uegnet for økologisk drift. Flere av de redskaper som er mest «på moten» i dag, er konstruert med tanke på at en ikke pløyer. Inntil noe annet er bevist, må en imidlertid si at riktig pløying er avgjørende for å holde rotugraset i sjakk.
Med tanke på ugrasregulering, er det viktig å velge riktig type harv og tinder for å få jevn sådybde og dermed jevn bestand av kulturvekster som kan konkurrere med ugraset. Kravet om jevn arbeidsdybde gjelder også arbeidsorgan og harver som skal brukes til brakking. Når en harver før såing med vanlig kornsåmaskin, skal harva normalt bare lage et løslag på toppen som er så dypt som en skal så. Sålabbene skal m.a.o. gå på harvbunnen. Det betyr at en skal ha et løslag på ca. 5 cm (3-4 cm etter tromling). Dyp harving gir lett for dyp såing, og dette kan gi dårlig oppspiring. På den andre siden er det uheldig å så for grunt, fordi da kan det bli for tørt for frøene til å spire, samtidig som det er større risiko for å rive opp frøene ved første ugrasharving. Er det mye rotugras, kan det være aktuelt med flere gangers harving, da dypere enn for såbed og med opphold mellom hver harving, slik at formeringsorganene dels tørker ut, og dels svekkes ved at de spirer og ødelegges på nytt. Gamle forsøk (harving med hest) viser reduksjon i kvekemengda med flere gangers vårharving. I økologisk drift bør en bestandig tenke jordpakking i forbindelse med jordarbeiding. En bør derfor ikke velge alternativer som er for tunge, og som krever stor traktor. En bør kjøre med brede dekk og/eller tvillinghjul med lavt lufttrykk, og fokusere spesielt på hjulene foran på traktoren dersom lasteren sitter på.
Harv var tidligere en generell betegnelse på de redskapene som ble brukt for å lage et såbed etter pløying. I dag har harvene fått et utvidet bruksområde, og overgangen mellom de forskjellige typene av harver er ganske glidende. Noen er bevisst laget universelle, mens andre er formet for helt spesielle oppgaver. Det vi kan klassifisere som harver, kan brukes til å lage såbed, løse opp og lage såbed i upløyd jord, regulere ugras og for innblanding av halm, planterester og husdyrgjødsel.
Skrapere er normalt flattstål som er sveiset fast under understrømningsplankene på slodder. De er montert på skrå i forhold til kjøreretningen slik at de bearbeider et relativt bredt belte. Når skrapere monteres på flere sloddplanker vil de gi en jevn harvbunn, og dersom de er høye nok, kan en så direkte etter slodden. På steinholdig jord er de mindre egnet.
Tindeharver er de eldste harvene vi har, og de er fortsatt aktuelle. De produseres med flere typer tinder, og tindene kan ha mer eller mindre kraftig konstruksjon etter hvilke oppgaver harva skal løse.
Stive tinder
Våronnharver med rette og krokete, stive tinder produseres knapt nok i dag, men de brukes på kraftuttaksdrevne harver som rotorharv og lett fres. Stive tinder er best egnet for grunn harving, for eksempel til korn, i områder med stiv jord (leirjord), og for dypere harving med de kraftuttaksdrevne harvene.
Stive tinder med og uten utløsere brukes til dypharver og grubbere. Stive tinder sikrer jevn harvedybde. Tinder med utløsere vil virke som en stiv tind inntil utløsingskrafta overstiges. Når tinden løser ut vil arbeidsdybden reduseres kraftig på den enkelte tinden. Tindene kan utstyres med gåsefotskjær slik at en får full gjennomskjæring.
Fjærende tinder
Konstruksjoner med fjærende tinder med utløsere ligner mye på stive tinder med utløsere, men tinden er i dette tilfelle laget av fjærstål. Den får derfor en vibrerende gange og dette reduserer faren for subbing i jord med mye planterester (figur 3.17).

C-tinden kom tidlig i bruk. Årsaken var at denne typen er enkel å produsere og man ønsket en fjærende tind. Når C-tinden møter motstand, vil den rette seg ut. På grunn av sin spesielle form, vil arbeidsdybden reduseres. Figur 3.18a viser dette. Den heltrukne streken viser formen på tinden ubelastet. Den stiplede streken viser tinden belastet. Figuren viser hvordan arbeidsdybden reduseres etter hver som motstanden øker. Denne egenskapen har sine negative og positive sider. Med henblikk på å harve for å få en jevn harvbunn og dermed jevn sådybde, er tinden dårlig, for når harva eller deler av harva kommer til harde områder, blir arbeidsdybden reduseret. Slik sett passer den bare for lettere jord. Imidlertid vil denne tindetypen være fordelaktig når en harver vollpløgsle. Under slike forhold vil tinder som holder relativt jevn arbeidsdybde lett kunne rive opp torv, mens C-tinden, som gir lettere etter, river opp mindre torv. For å få jevn arbeidsdybde må en da harve flere ganger.
C-tinden er knapt nok i produksjon i dag.
S-tinden dominerer på universalharver i dag. Grunnen er at den har en vibrerende gange og gir en relativt jevn arbeidsdybde. Figur 3.18b viser dette. Den heltrukne linja viser tinden ubelastet. Den stiplete linja viser tinden under belastning. Formen og stillingen på tinden gjør at arbeidsdybden varierer lite selv om motstanden varierer mye. Dersom S-tindharva er utstyrt med vridbare brander, kan en endre egenskapene til tinden. Figur 3.18c viser dette. Når branden vris, får S-tinden tilnærmet de samme egenskapene som C-tinden og egner seg bedre til vollpløgsle.
Brede S-tinder har i utgangspunktet samme egenskapene som universaltypen. Forskjellen ligger i at tindekonstruksjonen er kraftigere og kan utstyres med brede spisser og gåsefotskjær. De brukes for dypharver. De har en vibrerende gange, og subber mindre enn stive tinder med utløsere (figur 3.19a).
Smale S-formede tinder er spesialtinder beregnet på grunn harving, dvs. harver for å lage såbed. Tinden har i hovedsak samme konstruksjon som universaltinden, men den er smalere. Den delen som spissen er festet til, har en vinkel mot jorda som er nær 90o (figur 3.19d). Dette er fordi tinden har mindre behov for jordsøking, og vil ikke transportere rå jordklump opp til overflaten. I forhold til kraften den utsettes for, er den relativt stiv. En kan si at den kombinerer egenskapene til den rette tinden og S-tinden.
Kraftige Q-formede tinder lages ofte av kraftig, firkantet fjærstål. De er bøyd i en dobbel sløyfe og kan av utseende minne litt om en Q. Tinden har i hovedsak samme egenskaper som en kraftig S-tind, og brukes derfor til dypharver (figur 3.19b).

Lette Q-formede tinder lages vanligvis av rundt fjærstål og har en til tre sløyfer. Tinden er brukt til vanlige harver, men benyttes mest til etterharver på såmaskiner og andre harver. Den er beregnet på grunn bearbeiding (figur 3.19c). Tilsvarende tind brukes på noen ugrasharver.
Harver for grunn bearbeiding (såbedsharver)
Grunn bearbeiding vil si en harvedybde på under 6 cm. I praksis betyr det at harvene kun er beregnet på å lage såbed for korn og småfrø. Ofte kalles disse for såbedsharver. Harvene er vanligvis utstyrt med flere dybdehjul og/eller ribbetromler som sikrer eksakt dybde. Tindene er vanligvis smale S-tinder. Se figur 3.19d. Tindeavstanden er under 10 cm, vanligvis 5 til 7 cm. Selv om harvene har 4-5 brander, blir det relativt liten avstand mellom tindene og harvene har derfor lett for å subbe dersom det er mye ugras eller planterester. Av den grunn passer de mindre godt i økologisk drift. Fordelen med harvene er at de lager et godt såbed i én operasjon. Mest aktuelle er de på stivere jordarter og jord uten stein.
Harver for middels dyp bearbeiding (fjærharver)
Middels dyp bearbeiding vil si en harvedybde på fra 3-4 til 15 cm. Disse harvene kan derfor brukes til flere oppgaver i våronna. Vi skiller mellom C-tindharv og S-tindharv. Harvene har normalt en tindeavstand på 9-11 cm, og tindene er festet på 3 eller 4 brander.
C-tindharva var den opprinnelige fjærharva som dominerte for 50 år siden, og som fortsatt finnes i bruk. Fjærharva er utstyrt med faste meier eller dybdehjul. Dybden reguleres ved å vri brandene. Ulempen med dette er at en samtidig endrer vinkelen mellom spissene og bakken. C-tindens konstruksjon gjør at de egner seg best på lette jordarter og på vollpløgsle. C-tindharva er i de fleste tilfeller for myk til å gi tilfredsstillende harvedybde for poteter og andre vekster som krever dyp bearbeiding. Kvernelands Kultisvans som ble solgt i stort antall hadde stivere tinder og kunne utstyres med sloddplanke og ribbetrommel.
S-tindharva (figur 3.20a) er den dominerende våronnharva der det pløyes. Den har vanligvis dybdehjul, men det finnes også harver som bæres av ribbetromler. Den siste typen egner seg ikke på steinholdig jord fordi ribbetromlene vil løfte opp harva og redusere arbeidsdybden når de passerer steiner. Noen av harvene leveres med vribare brander. Det gjør harva mer universell, og harvene kan derfor brukes på stiv jord og på vollpløgsle. I økologisk drift bør en velge denne typen. Spissene på S-tindharvene er vanligvis 3-4 cm brede. Når de er så smale, vil de i liten grad transportere opp rå jord som blir liggende på overflaten og tørke til klump. Denne typen S-tindharver kan harve grunt dvs. 3-4 cm og ned til 15 cm. Flere har et bredt midtfelt og oppslagbare sidefelt. Ved grunn harving kan en bruke hele harva, mens en kan bruke bare midtseksjonen ved dyp harving dersom det skorter på traktorens trekkraft. Disse harvene har for liten tindeavstand til å kunne brukes til brakking på halmstubb eller der det er mye røtter og jordstengler fra rotugras. Harvene kan i de fleste tilfeller levers med sloddplanke, ribbetrommel eller etterharv.
Harver for dyp bearbeiding - dypharv (stubbharver)
Dyp bearbeiding vil si en harvedybde på 20-25 cm (opp mot pløyedybde). Ofte kalles de for stubbharver (figur 3.21b), fordi de er meget godt egnet til harving i upløyd jord, dvs til brakking høst og vår. I tillegg er disse harvene meget godt egnet for harving til poteter og til harving på nybrott. Disse harvene leveres med stive tinder, stive tinder med utløsere eller fjærende tinder. Fjærende tinder er i de fleste tilfeller fordelaktig fordi de på grunn av vibrasjonen i tinden lettere kvitter seg med planterester og røtter, og har derfor ikke så lett for å bli blokkert av planterester. Harvene har normalt en tindeavstand på 20-25 cm. De har to eller tre brander. Med tanke på blokkering er det gunstig med stor avstand mellom tindene både i lengderetning og sideveis. Skal en harve meget dypt, bør den største tindeavstanden velges. For harveresultatet er det imidlertid en fordel at tindeavstanden ikke er mer enn 20 cm. En bør derfor velge harver med tre brander. Det er dessuten en fordel om det er enkelt å endre tindeavstanden.
Harvene har normalt spisser som er 5-12 cm brede. De kan utstyres med vridde spisser som bedrer jordblandingen. De kan også utstyres med gåsefotskjær som gir full gjennomskjæring. Et firma kan levere spissfester med hurtigkopling av spissen. Med dette er det fort gjort å skifte mellom vanlige spisser og gåsefotskjær. Med tanke på brakking, er disse harvene dårligere egnet enn en skikkelig skålharv eller en roterende harv. Stubbharva river opp røttene, men deler ikke opp røttene i tilstrekkelig grad. Dersom en har slik harv, er den absolutt brukbar, men den må kjøres flere ganger. I de senere årene har det kommet en type som kalles skålgrubber (figur 3.21f). Den har kraftige tinder med eller uten utløsere og er utstyrt med kraftige gåsefotskjær og spisser. Harva har en skålrekke bak tindene som skal jevne ut jorda etter tindene, derfor navnet. Den er også utstyrt med en eller annen type pakkevals bakerst som skal pakke sammen og jevne jordoverflaten. Dette er redskaper som kan brukes høst og/eller vår for bearbeiding av upløyd jord. Harva er meget tung og krever stor traktor, og derfor lite aktuell for økologisk drift.
Harver for meget dyp bearbeiding (grubbere)
Meget dyp bearbeiding vil si en harvedybde dypere enn vanlig pløyedybde. De kalles vanligvis for grubbere og kan være vanskelige å skille fra dypharver. Grubbere kan brukes for å løse opp pakkeskader i jorda og dessuten til brakking. Dypløsning bør foretas med én kjøring tidlig på høsten når det er tørt. Da gjør en minst mulig skade, og jorda får «satt» seg til neste vår.
Skålharver
Skålharver (figur 3.21c og d) ble tidligere brukt til harving av vollpløgsle. Til dette arbeidet ble det brukt lette typer. I dag har vi flere alternativer. Ut i fra bruksegenskaper og tyngde kan vi skille mellom tre hovedtyper, vist i tabell 3.6.
Tabell 3.6 Skålharvtyper
| Skåldiameter | Vekt per meter arbeidsbredde uten pakkevalser |
|
| Lett skålharv | < 45 cm | < 250 kg |
| Middels tung skålharv | 45-55 cm | 250-500 kg |
| Tung skålharv | > 55 cm | > 500 kg |
Lett skålharv er først og fremst beregnet på å lage såbed på pløyd mark, spesielt vollpløgsle. Til å bekjempe rotugras blir den normalt for lett, dersom man da ikke løser opp på forhånd med en kraftig tindeharv eller bruker den på de letteste jordtypene uten stein.
Middels tung skålharv er en harv som egner seg godt til stubbharving, dersom jorda ikke er for hard og tørr, og hvor halmen er fjernet. Den er meget godt egnet til såbedstilberedning dersom en kan begrense arbeidsdybden der det er løst, med hjul eller valse.
Tung skålharv er et utmerket redskap når en skal tyne rotugras. Den skjærer seg ned og kutter formeringsrøtter og jordstengler i hard jord, også der det er mye halm på overflaten. Det er viktig at den har gode nok dybdehjul eller pakkevalse bak. Den tunge typen er dyr i innkjøp, men er mer robust mot slitasje.
Skålharvene kan som oftest belastes mer. Nyere typer er ofte utstyrt med pakkevalse etter skålseksjonene. I praksis kan derfor sum vekt per meter arbeidsbredde bli vesentlig større enn angitt i tabell 3.6, og typene vil gli over i hverandre. De fleste kan utstyres med slette eller takkete skåler. De slette gir jevnere gjennomskjæring og dermed bedre oppdeling av ugras, mens de takkete trenger noe lettere ned i bakken når det er hardt.
Skålharva skilles fra tindeharva ved at den skjærer et vertikalt snitt og at den blander jorda meget godt. Det er derfor tre områder den er særlig godt egnet til: Tilberedning av såbed på ompløyd voll, brakking mot rotugras og nedmolding av husdyrgjødsel. Problemet med skålharva er at det i hovedsak er vekt, stein i jorda og jordhardhet som bestemmer hvor dypt den går. Den søker ikke nedover slik som tindeharva. Har harva gode nok dybdehjul eller pakkevals bak, kan en regulere maksimumsdybden til harva. En skal imidlertid være klar over at når harva belastes for å gå dypere, økes også slitasjen.
Tidligere satt alle skålene i hver seksjon på en felles aksel. Harvene hadde da en X-form (skålene sitter på fire aksler) eller en V-form (skålene sitter på to aksler som dannet en V-form på tvers av kjøreretningen) (figur 3.21c). Skålenes vinkel på kjøreretningen kunne endres ved å endre vinkelen på akslene i forhold til kjøreretningen. På X-formen vil de to framre skålrekkene kaste jorda utover og den bakre innover igjen. På disse harvene er det viktig å regulere arbeidsdybden på framre og bakre rekke slik at like mye jord kastes innover som det er kastet ut. Går den bakre rekka for grunt, blir det en forsenkning i overflata midt bak harva, og går den for dypt, blir det mye jord midt bak harva. V-formen vil på samme måte kunne transportere jord sideveis, og kjører en tilbake langs draget en kjørte fram, kan det bli forhøyning eller forsenkning avhengig av om den bakre skålseksjonen går for dypt eller for grunt. I de siste årene har det kommet skålharver hvor hver skål sitter på en egen opphengsarm med fast eller stillbar vinkel i forhold til kjøreretningen (figur 3.21d). Armen har som oftest fjæring, slik at den enkelte skåla kan gå opp over en stein uten at hele skålseksjonen løftes. Disse er noe enklere når det gjelder å få lik arbeidsdybde foran og bak fordi det er kort avstand mellom seksjonene. Denne typen skålharv leveres enten for sleping eller for montering på trepunkten. De som er montert på trepunkten, krever en traktor med stor løftekraft, og den må ha stor vekt foran.
Kraftuttaksdrevne harver
Ved å drive jordarbeidingsorganet med kraftuttaket, reduseres behovet for trekkraft vesentlig. Dette skulle i utgangspunktet bety at en kunne bruke en vesentlig lettere traktor, men fordi disse harvene har stor vekt, må traktoren likevel være stor og dermed tung, for å kunne løfte dem. Ved å drive arbeidsorganene med kraftuttaket vil en kunne tilpasse bearbeidingsintensiteten ved å kombinere turtall og framdriftshastighet. Dette er i utgangspunktet en fordel, men i praksis har en ofte sett at bearbeidingsintensiteten blir for voldsom, og at jordstrukturen blir ødelagt. Det er ikke noe stort problem å få slike redskaper til å arbeide dypt. Dette kan bety at tindene kommer ned i fuktig jord som er enda mindre sterk mot mekanisk påkjenning. Kraftuttaksdrevne harver må derfor ha hjul eller være utstyrt med valse bak som kan begrense arbeidsdybden. Det er viktig å forstå denne sammenhengen, slik at de kraftuttaksdrevne harvene utfører et positivt arbeide og ikke ødelegger jorda.
Det er to typer som brukes: fresen, hvor alle arbeidsorganene sitter på en horisontal aksel på tvers av kjøreretningen, og rotorharva, hvor to og to arbeidsorgan sitter på flere (mange) loddrettstilte aksler.
Jordfresere (figur 3.20b) opptrer i flere varianter og fasonger. Drifta kommer vanligvis inn på midten av akselen, men den kan også komme inn på en av sidene. Hastigheten som et arbeidsorgan treffer jordpartiklene med, er en kombinasjon av framdriftshastighet og arbeidsorganets periferihastighet. Hastigheten, og dermed påkjenningen på jorda, er størst der arbeidsorganene går ned i bakken og minst loddrett under akselen. For at arbeidsorganene i det hele tatt skal gjøre noe arbeid loddrett under akslingen, må periferihastigheten være større enn framdriftshastigheten. Loddrett under akselen er den reelle hastigheten til arbeidsorganet mot jorda periferihastigheten minus framdriftshastigheten. Der arbeidsorganet går ned i jorda, vil den reelle hastigheten være omtrent lik periferihastigheten, og dersom en kjører meget dypt, det vi si en dybde tilsvarende radien på arbeidsorganet, vil den reelle hastigheten være større enn periferihastigheten. Det er derfor viktig å avpasse framdriftshastighet, rotorhastighet og dybde, slik at jorda ikke blir ødelagt. Arbeidsorganene har normalt ikke fjæring og de treffer stein med større hastighet enn trukne harver. Dette gjør at de må være meget robuste for å kunne benyttes på steinholdig jord. I mange tilfeller opplever en at arbeidsorgan knekker og/ eller bøyer seg, og at festebolter klippes av, spesielt dersom de er dårlig tiltrukket. Uansett kan en få store vedlikeholdskostnader på hard og steinholdig jord. Fresen kan brukes til å lage såbed og til bekjempelse av rotugras. Den vil også være meget godt egnet til å blande inn planterester og husdyrgjødsel. Arbeidsorganene kan variere fra rette kniver (rundtstål, firkantstål eller flattstål), til J-formede kniver eller L-formede kniver. De rette typene kan brukes til de fleste oppgaver, mens de L-formede er best når en ønsker å dele opp røtter. I forbindelse med bekjempelse av rotugras, vil J- og L-formede stål være gunstigst.
Også for rotorharva (figur 3.20c) vil hastigheten som arbeidsorganet treffer jordpartiklene med, være avhengig av framdriftshastighet og periferihastighet på arbeidsorganene. Her er kanskje bildet noe enklere. Det arbeidsorganet som er på tur framover vil oppnå en maksimalhastighet som er summen av framdriftshastighet og periferihastighet. Det som er på tur bakover, vil ha en hastighet som er periferihastighet minus framdriftshastighet. Dersom arbeidsorganet har en hastighet lik framdriftshastigheten når det går bakover, vil det ha en hastighet lik tre ganger framdriftshastigheten når det går framover. En kan derfor ikke oppnå stor arbeidskapasitet med dette redskapet dersom en ikke skal ødelegge jord. Når det er sagt, vil redskapet være effektivt i kampen mot rotugras, stort sett på linje med en fres med rette tinder. Rotorharva er også effektiv når den koples sammen med ei såmaskin. På slike kombiredskaper sitter såmaskina festet på rotorharva. Når en harver og sår samtidig, passer det å kjøre med en hastighet på 4-7 km per time. Da blir systemet effektivt fordi en gjør to operasjoner samtidig, og harva øderlegger ikke jorda. Positivt med rotorharva er også at en ikke trenger å kjøre mer enn en tur over jordet, mens en kanskje må kjøre to turer med ei tindeharv. Med pakkevalse bak, er en sikret meget jevn harvdybde.

Vedlikehold av harver
Skal en harve til jevn dybde, må alle organene på harva gå like dypt. Dersom tinder strekkes slik at spissen går grunnere, må de skiftes ut. Når noen av spissene må skiftes ut (snus), må alle skiftes slik at de arbeider like dypt. Slitelengden på en spiss er nesten like lang som harvedybden, og da er det innlysende at resultatet blir dårlig dersom det står en ny og en nesten nedslitt spiss ved siden av hverandre. Ta gjerne vare på halvslitte spisser, de kan settes på dersom en skulle være uheldig å miste en spiss når alle er halvslitte. Om en mister eller bøyer et arbeidsorgan på lette freser eller rotorharver, og må skifte dette, bør en kutte det nye arbeidsorganet, hvis de andre er halvslitte slik at alle arbeider like dypt.
Kjøremønster ved harving
Mye av det som er sagt om kjøring på vendeteiger osv. under slodding, gjelder også når en harver, se kapittel om Harving. Harver med tinder kan være meget effektive til å dra opp kveke og spre den utover. Det kan være en fordel å løfte opp harva på vendeteigene. Dersom en kjører teiger og unnlater å løfte opp harva på vendeteigene, vil jorda der bli pulverisert, og det blir dårlig vekst. Veksten på vendeteigene er spesielt viktig for å kunne konkurrere med ugraset.
Har en jevn pløgsle, god slodd og god harv, vil det som oftest være tilstrekkelig med én gangs harving til korn og gjenlegg (dersom en ikke skal bekjempe rotugras). Det kan imidlertid være områder på jordet hvor det er større behov for harving. Da kan en harve disse områdene først, og deretter harve hele stykket. Vekster med kraftige røtter som gulrot krever djupere løsning av jorda.
Jordarbeiding, sammen med konkurransen mellom kulturplante og ugras, er på mange måter fundamentet for ugraskampen. Dette gjelder innenfor enhver driftsform, men i økologisk landbruk er en optimalt utformet jordarbeiding spesielt viktig. Hvilke vekster man dyrker er avgjørende for når og hvor lenge man kan gjennomføre jordarbeiding. Det er derfor naturlig å se på jordarbeidinga som en integrert del av plantedyrkinga på en gård. Følgelig er det vanskelig å skille effekter av jordarbeidinga fra det man gjerne kaller en vekstskifteeffekt. For jordarbeiding med det mål å kontrollere ugras vil kunnskap om ugrasets biologi, og hvordan biologien blir påvirket av andre faktorer, for eksempel pløyedybde, være en helt nødvendig plattform å bygge jordarbeidingsstrategien på. Noe av målet med dette avsnittet er derfor å trekke tråder mellom jordarbeidinga og ugrasets biologi.
Jordarbeiding har to forskjellige virkninger på ugraset:
Frøugras og jordarbeiding
Jordarbeiding stimulerer den biologiske aktiviteten i jorda og påvirker i stor grad også ugrasfrøene. Jordarbeiding medfører generelt en sterk økning i antall frø som spirer (men variasjon mellom ugrasarter finnes) og dermed også antall oppkomne planter. Effekten av jordarbeiding er avhengig av bl.a. jordfuktighet. Flere frø spirer hvis jorda er fuktig. Mange av de frøene som spirer i forbindelse med jordarbeidinga, vil av ulike årsaker dø den første tida og dermed aldri konkurrere med kulturplantene (se tabellene 3.7 og 3.8).
Tabell 3.7 Effekt av jordarbeiding på frøbanken1
| Behandling | Forandring av frøbank i % av hva den var i begynnelsen av vekstsesongen | ||
| Total nedgang i frøbanken | % spirte planter | % frø som døde | |
| Ubehandla | 22 | 1 | 21 |
| Jordarbeiding 2x | 30 | 7 | 23 |
| Jordarbeiding 4x | 36 | 9 | 27 |
| «Helbrakk» | 50 | - | - |
1Etter Håkansson 2002
Tabell 3.8 Antall spirende frø fra ulike jorddyp (8 dager etter jordarbeiding)1
| Dyp, cm | Antall spirende frø per m2 |
| 0-5 | 2437 |
| 5-10 | 69 |
| >10 | 0 |
1Mest åkersennep, pengeurt, meldestokk, rødtvetann. Etter Håkansson 2002
Etablerte planter av ettårige ugras blir lett skadet om de blir dekket skikkelig av jord. I fuktig jord vil de komme seg raskere igjen enn i tørr jord, og dette gjelder spesielt om de er ufullstendig tildekket. Spirende planter vil være mest ømfintlige rett etter oppspiring. Dessuten vil planter av småfrøete arter, fordi de har mindre reservenæring, være mer ømfintlige for forstyrrelse enn storfrøete arter.
Hvilken effekt man kan forvente av jordarbeiding på sommer- og vinterettårige arter kan man lese ut fra figur 2.18. Frøa hos disse artene kan spire og utvikle seg om våren selv uten jordarbeiding, men jordarbeiding vil ofte øke spiringen betydelig. Spireviljen til sommerettårige arter avtar sterkt utover forsommeren (se omtale av «dormans» i kapitlet Skadegjørernes livsstrategier), og jordarbeiding utover i vekstsesongen vil derfor i mindre grad påvirke disse artene. På den annen side vil vinterettårige arter være spirevillige utover hele vekstsesongen og derfor kunne spire også etter jordarbeiding.
Flerårige ugras og jordarbeiding
Også flerårige arter produserer frø, og når disse spirer, vil situasjonen naturligvis være den samme som for ettårige arter, men når vegetative organer (formeringsrøtter, jordstengler m.m.) etter hvert blir dannet, vil potensialet for å overleve jordarbeiding ofte øke betydelig. På den annen side vil visse arter som løvetann, høymole, sølvbunke, krypsoleie, stormaure og stornesle fremdeles være følsomme. Andre flerårige arter som åkertistel, åkerdylle og kveke, er langt mer robuste mot jordarbeiding.
Jordarbeiding vektlegges tungt i forbindelse med ugrasbekjemping, særlig bekjemping av arter med krypende formeringsorgan som røtter eller jordstengler. Disse artene er viktige ugras i ettårige åkervekster fordi de er vanskelige å ta både med pløying og ved tillaging av såbed. Blant disse artene kan kveke, åkerdylle, storkvein, vass-slirekne, med relativt grunt utløpersystem nevnes, mens hestehov og åkertistel har dypere utløpere (for enkelte av artene, se figur 2.23).
For alle disse artene må jordarbeidinga gjennomføres på en skikkelig måte for at denne skal være effektiv. Kunnskap om ugrasets biologi, jfr. kapitlet Skadegjørernes livsstrategier, vil være en helt nødvendig plattform å bygge jordarbeidingsstrategien på. Kunnskapen om biologi er viktig både for å avgjøre hvilke redskapstype som skal benyttes, når på året og hvor ofte tiltaket skal settes inn.
Følgende faktorer er viktige å ta hensyn til ved utarbeidelse av effektive jordarbeidingsstrategier:
Viktigheten av dyp pløying mht effekten på rotugras er godt dokumentert både i eldre og nyere forsøk. I et norsk forsøk har totalmengde rotugras blitt mer enn halvert ved dyp sammenlignet med grunn pløying (se tabell 3.9).
Tabell 3.9 Tørrstoffmengde (gram per m2) av ulike rotugras i forsøk på Kvithamar (Trøndelag)
og Apelsvoll (Oppland) som følge av grunn (15 cm) og dyp (25 cm) pløying1.
| Lokalitet | Pløyedybde | Totalt2 | Åkertistel | Åkerdylle | Kveke | Hestehov |
| Kvithamar | Grunn | 38,7 | 6,6 | 0,4 | 19,8 | 11,8 |
| Dyp | 23,3 | 3,3 | 0,2 | 15,7 | 3,8 | |
| Apelsvoll | Grunn | 50,5 | 7,3 | 38,5 | 2,9 | 0 |
| Dyp | 16,9 | 0,7 | 11,3 | 2,6 | 0 |
1Etter Brandsæter et al. 2005. 2Også andre ugras enne de spesifiserte inngår i «Totalt».
Ved jordarbeiding til vanlig tid om våren er målet å lage et så- eller plantebed med færrest mulig spirte ugrasplanter. Jordarbeidet skal ikke gjøres dypere enn det som er nødvendig for å lage et godt såbed, men samtidig vil grunn jordarbeiding ha liten effekt på flerårige ugras. Jordarbeiding og såing eller planting kan gjøres på samme tid, gjerne i samme operasjon. Figur 3.36 viser hvor viktig det er at kulturplanten spirer raskest mulig i forhold til ugraset. Hvis vi i denne omgang ser bort fra faren for erosjon og utvasking, anbefales høstjordarbeiding fordi dette gir jevnest såbed, og man unngår omfattende flytting av jord om våren.
Frøugras
Flere undersøkelser har konkludert med at utsatt såtid om våren har en gunstig effekt på ugrassituasjonen. Blant annet har en flerårig forsøkserie i konvensjonelt dyrket korn i Norge vist at antall ugrasplanter avtar med utsatt såtid (figur 3.23). Dette har to årsaker:
Selv om utsatt såtid medfører mindre oppspiring av ugras, er ikke metoden uten ulemper. Kulturplanter som krever relativt lang vekstsesong, blir mer avhengig av værforholdene utover sensommeren og høsten for å bli høsteklar. En annen potensiell ulempe kan være manglende spirefukt om våren. Den omtalte forsøkserien har helt klart vist at utsatt såtid i mange år også betyr nedsatt avling (figur 3.24).
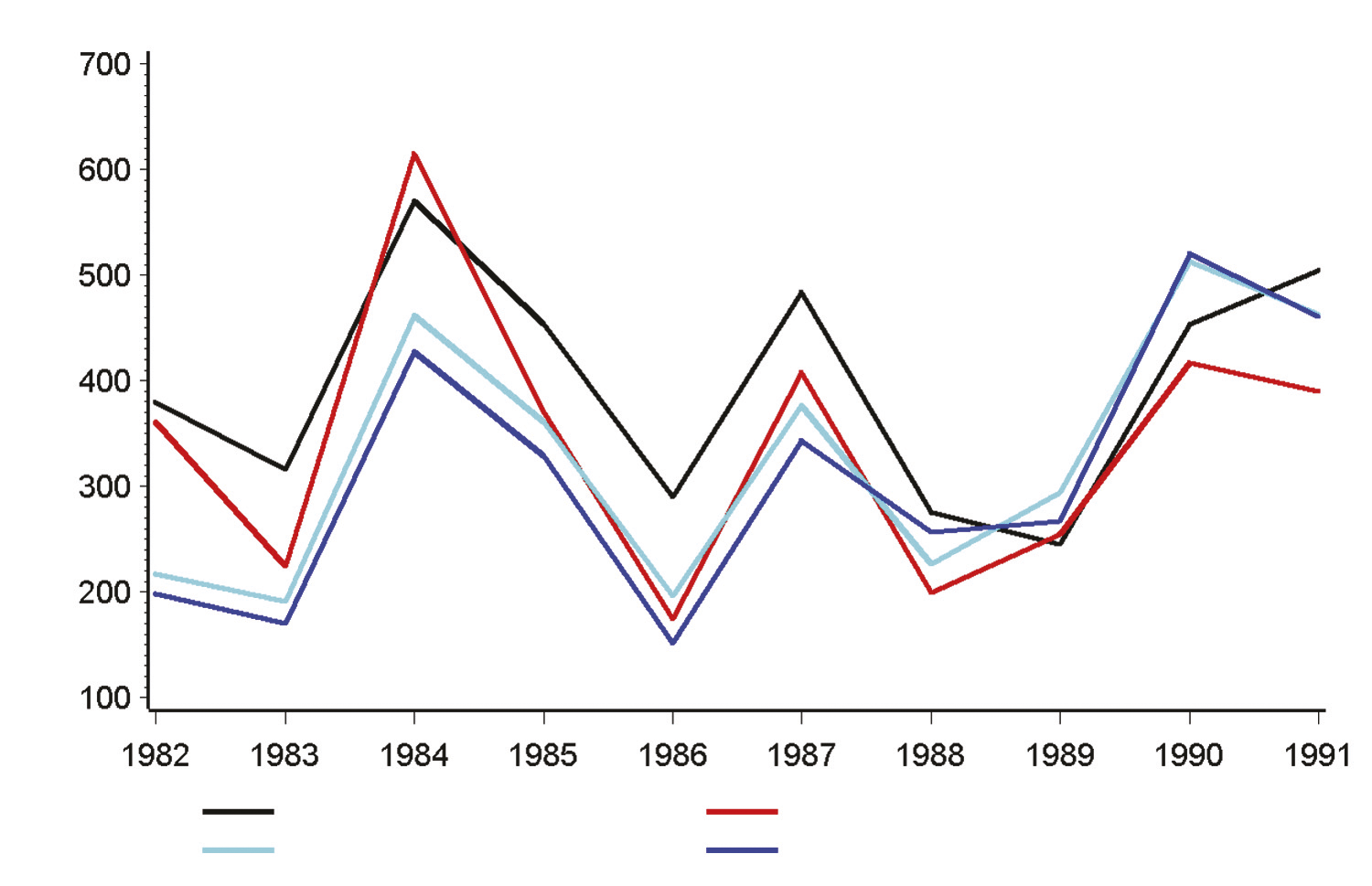
Utilstrekkelig spirefukt har i alle fall tidligere vært viktig på Østlandet hvor forsommertørke tradisjonelt har vært et problem. Når det gjelder utsatt såtid, må hver enkelt bonde gjøre sin egen vurdering om metoden er aktuell for produksjonene på gården. Ved økologisk dyrking kan et aspekt være at utsatt såtid gir varmere jord ved såing, enn såing ved vanlig tidspunkt. Dette kan være et pluss for mineralisering og dermed tilgangen på næringsstoffer fra organisk materiale. Det kan her legges til at vårkornet i eldre tider i gjennomsnitt ble sådd et par uker senere enn i dag, i en del tilfeller enda senere.
Flerårig ugras
Jordarbeiding om våren og forsommeren har god effekt på flerårige ugras fordi plantene på denne tida er i god vekst, og knoppene på røtter og rhizomer er ikke-dormante/spiretrege. Man bør harve, frese eller lignende slik at de vegetative formeringsorganer blir delt opp mest mulig og knoppene stimulert til å sette lysskudd. Behandlinga bør gjentas, slik at man får «sulta ut» formeringsorganene mest mulig. Hvor lang tid det bør være mellom hver behandling er blant annet avhengig av hvilke ugrasarter som skal bekjempes. Man skal iallfall ikke vente lengre enn at ugraset har nådd stadiet for tørrstoffminimum i røtter eller jordstengler (se figur 2.21). Generelt vil ugraset utarmes noe raskere hvis man jordarbeider før plantene når stadiet for tørrstoffminimum. Figur 2.26 viser f. eks. at behandling av kveke på 1-2-bladstadiet fører til raskere utsvelting enn behandling på 3-4-bladstadiet. For åkertistel har forsøk i USA, hvor det ble harvet 15 cm dypt med 1-6 ukers intervaller, vist at harving hver 3. uke gav raskest effekt. Hvor lang brakkingsperioden bør være for å gi god kontroll av ulike ugras, vet vi for lite om i dag.
Selv om det ikke er lett å konkludere hvilken redskapstype som er best for å tyne ugraset, kan vi slå fast at redskap som kutter opp røtter eller jordstengler best mulig, vil være å foretrekke. Flere rotbiter vil gi flere lysskudd, og det betyr at plantenes opplagsnæring raskere blir brukt opp. Et annet viktig aspekt er om redskapen som benyttes, når ned til røttene. Dette vil som regel ikke være noe problem for gruntvoksende arter som kveke og åkerdylle, men for åkertistelen kan situasjonen være annerledes fordi de horisontale formeringsrøttene ligger dypere, for det meste i sjiktet 15-30 cm. Har man mye åkertistel, kan det være aktuelt å starte jordarbeidinga med dyp pløying slik at redskapen når ned til disse røttene og får dratt dem opp. Det foregår for tida forsøk i Norge for å belyse dette nærmere.
Hvis man skal følge resultatene fra modellforsøk (se figurene 2.24 og 2.25), bør man etter å ha kuttet opp røtter eller jordstengler pløye disse lengst mulig ned. Lysskuddene vil da enten ha en redusert mulighet for å komme opp (ikke nok opplagsnæring), eller så vil lysskuddene bruke lang tid opp til overflata og dermed ha svært dårlig konkurranseevne. Hvis disse svake skuddene utsettes for sterk konkurranse fra en kultur, vil den kunne være nådestøtet for mange slike lysskudd. Denne metoden er kanskje mest aktuell i et år hvor man dyrker en grønngjødslingsvekst fordi denne tåler å bli sådd relativt sent.
Etter for eksempel en tidligkultur av grønnsaker eller poteter, eventuelt ved avslutning av ett eller flere engår (se tekstboks 3.6), kan man gjennomføre en kort eller lengre periode med brakking før man for eksempel sår høstkorn. En slik brakkingsperiode kan være effektiv mot flere problematiske flerårige ugras, herunder kveke, åkertistel og åkerdylle. I likhet med vårbrakk vil man ved midtsommerbrakk unngå hvileperiodene (dormansen) til disse artene.
Kveka kan gjøre seg sterkt gjeldende i eldre eng og forårsake stor skade i neste kultur i omløpet, spesielt ved økologisk drift. Ulike strategier som kan kontrollere kveka bedre ved avslutning av engperioden i et omløp er undersøkt. Forsøket ble gjennomført på sandjord i eng bestående av timotei og rødkløver med følgende behandlinger 3. engår eller våren etterpå:
Hvordan de ulike behandlingene påvirket kveka, ble registrert i bygg året etter. De alternative metodene reduserte tettheten av kveka sammenlignet med vanlig praksis. For behandling 3, 4 og 5 varierte antall kvekeskudd mellom 147 og 182 per kvadratmeter, men det var ingen statistisk sikker forskjell mellom disse behandlingae. Det var færrest kvekeskudd for behandling 1, midtsommerbrakken, hvor det bare var 27 skudd per kvadratmeter. Behandling 2, stubbarbeidet etter to høstinger av enga, lå i en mellomstilling , med 94 skudd per kvadratmeter. Også mht. byggavlinga ble det tallmessig beste resultatet oppnådd etter «midtsommerbrakk», men det ble ikke funnet statistisk sikre avlingsforskjeller mellom noen av behandlingene. (Etter Rouhiainen et al. 2003)
Hvis åkertistel eller andre ugras med dypt rotsystem er hovedproblemet i åkeren, kan det være aktuelt å pløye før jordarbeiding. En ulempe ved å pløye før brakkperioden kan være at det blir vanskelig å pløye etter avsluttet jordarbeiding. Som allerede nevnt, gjennomføres det nå forsøk for å belyse pløyetidspunkt i forhold til brakking, og vi vil derfor etter hvert kunne fortelle mer om denne teknikken.
Frøugras
Hovedtyngden av spirende frøugras om høsten vil være vinterettårige og toårige arter, men en del sommerettårige arter, spesielt ved tidlig såing, vil også spire. Det meste av de spirte sommerettårige plantene vil selvfølgelig dø i løpet av vinteren. Jordarbeiding om høsten vil stimulere ugraset til å spire, og en gjentatt behandling vil være med på å redusere frøbanken av disse artene. I forbindelse med dyrking av høstkorn vet vi at utsatt såtid vil redusere ugrasmengda ganske sterkt. Som om våren (vårbrakk) vil effekten av en slik utsatt såtid skyldes to ulike årsaker: (1) Man reduserer frøbanken i spiresjiktet ved gjentatt jordarbeiding, (2) Utover høsten blir en større del av frøene dormante, og vil ikke spire selv om man jordarbeider. Forsøk i høstkorn i Norge har vist klare effekter av utsatt såtid på ugrasutviklinga (tekstboks 3.7).
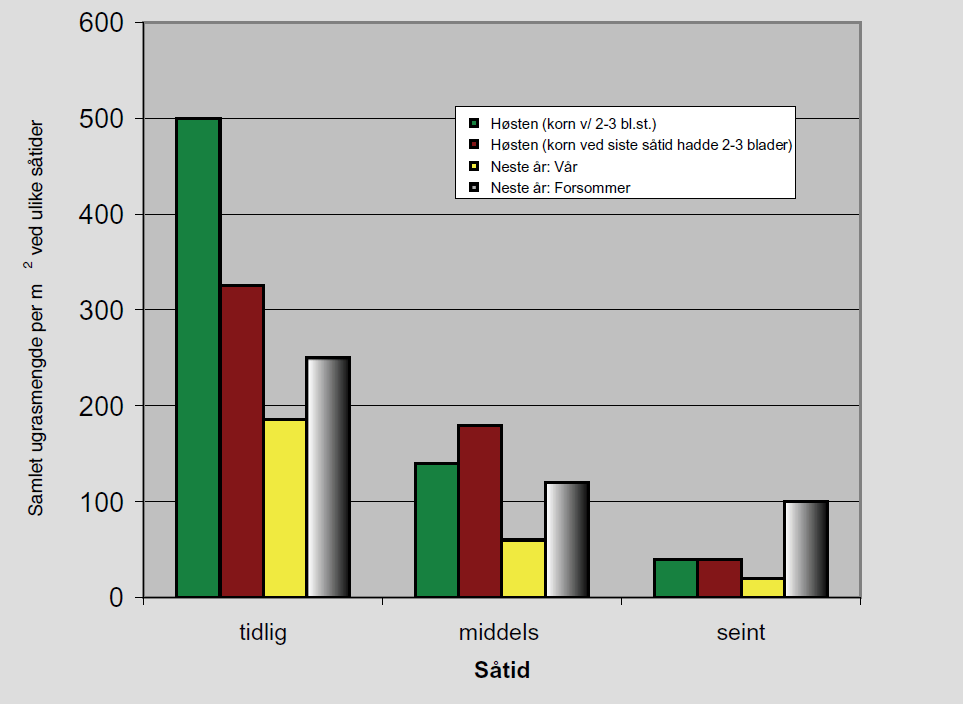
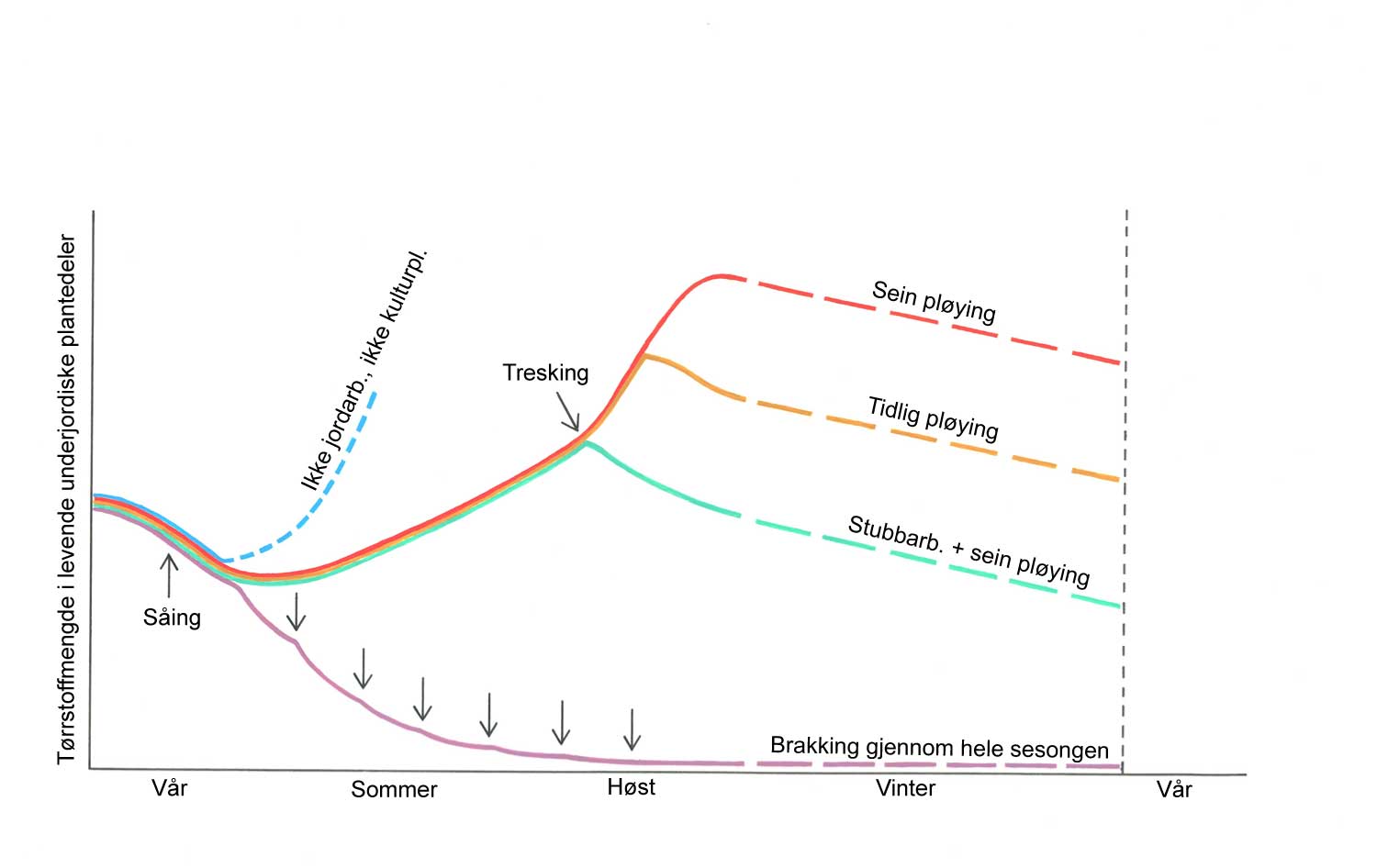
Hvilken effekt har så jordarbeiding på andre ugrasarter? Ikke alle ugras er så spire- og vekstvillige om høsten som kveka, og dette har konsekvenser for hvordan tiltakene bør utformes. Spesielt åkerdylla er spiretreg om høsten og en oppkapping av røttene på denne tida vil ikke bety at nye lysskudd blir dannet. Oppkapping av dyllerøttene kan i verste fall føre til oppformering av et stort antall nye planter i neste kulturvekst. Åkertistel er ikke så spiretreg som åkerdylla, men vi har noe av det samme fenomenet også for den. Ulike metoder for kontroll av flerårige ugras om høsten ble vurdert i et ganske omfattende svensk forsøk (tekstboks 3.8).
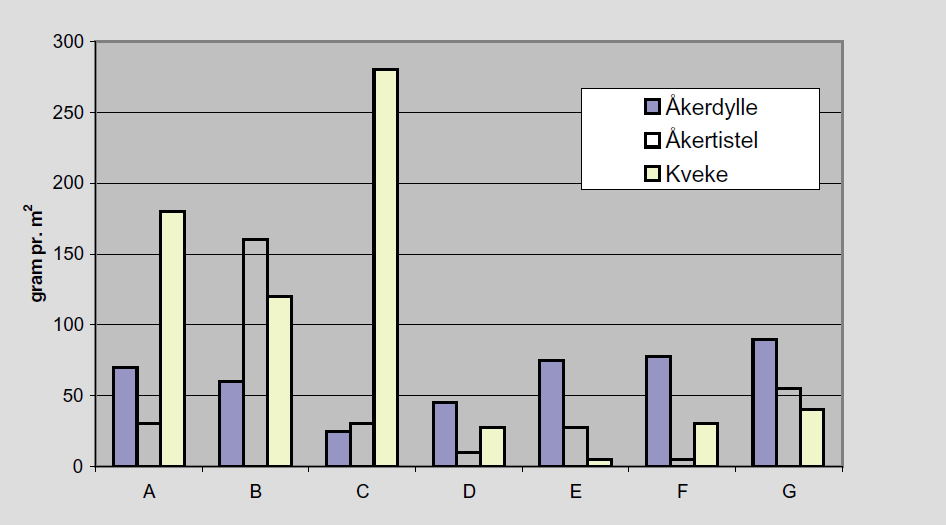
| Behandling | ||||||
| A | B | C | D | E | F1 | G |
| Tidlig pløying | Tidlig pløying | 0 | 0 | 0 | 0 | 0 |
| 0 | Harving | 0 | Grunn stubbarbeiding | Dyp stubbarbeiding | Dyp stubbarbeiding | Dyp stubbarbeiding |
| 0 | 0 | Sen pløying | Sen pløying | Sen pløying | Sen pløying | 0 |
1Som E, men bare hvert annet år. Som tabellen viser, har behandling A og C bestått av henholdsvis tidlig og sen pløying om høsten. Behandling B besto av tidlig pløying etterfulgt av harving. Behandling D og E besto av sen pløying etter hhv. grunt og dypt stubbarbeid. Behandling F var helt lik E, bortsett fra at den bare ble gjort annet hvert år. Behandling G besto av dyp stubbarbeiding og ingen pløying.
Grunn stubbarbeiding ble utført med spaknivharv med arbeidsdybde 5 cm. Til dyp stubbarbeiding ble det benyttet en stubbkultivator med arbeidsdybde 10 cm. Det ble pløyd ned til 25 cm. Forsøket ble utført under konvensjonelle forhold hvor også alle forsøksledd ble kombinert med ulike former for ugrassprøyting i tillegg til ubehandla kontroll. Resultatene er gjennomsnitt av de ulike sprøytebehandlingene og kontrollen. Selv om forsøket ikke ble gjort ved økologisk dyrking, er resultatene ganske illustrerende for hvilke tiltak som er mest effektive for kontroll av de ulike artene.
Med hensyn til kveke ser vi at lite jordarbeiding om høsten (A, B og C) gav mye kveke året etterpå. Verst blei situasjonen etter kun sen pløying (C). Da fikk kveka god anledning til å formere seg utover høsten. Behandlingene med stubbarbeiding (D, E, F og G) gav best resultat. Dette stemmer godt med gamle råd for kontroll av kveke, som sier at stubbarbeiding utover høsten kombinert med en avsluttende, gjerne dyp, pløying gir god kvekekontroll.
Ser vi «litt stort på det», kan vi for åkerdyllas vedkommende hevde at resultatene er omvendt av effekten på kveka. Dyp stubbarbeiding (E, F og G) gav dårligst kontroll av åkerdylla. Stikk motsatt av hva vi så for kveke, har det minst behandla leddet, dvs. bare sen pløying (C) gitt minst åkerdylle. Sterk oppkapping av røttene om høsten medførte derimot et økende problem året etter.
Utslagene for åkertistel er litt vanskeligere å oppsummere fordi det ikke er «signifikante» forskjeller mellom de fleste behandlinger, unntatt for behandling B. Det helt tydelige utslaget var altså at pløying først og harving etterpå gav sterk oppformering av åkertistelen. Dette skyldes sannsynligvis at pløyinga først har transportert en stor del av formeringsrøttene opp mot overflata der de så er blitt kuttet opp i småbiter ved harvinga etterpå. Rotbitene har så etablert nye planter neste år.
Den mest ytterliggående form for jordarbeiding vil være mekanisk brakking gjennom hele vekstsesongen (helbrakk). Undersøkelser har vist at dette kan gi om lag 50 % reduksjon i frøbanken. Effekten vil være størst under fuktige forhold. Frøhvile reduserer brakkingseffekten, noe som for eksempel vil gjelde sommerettårige arter utover i sesongen.
En lang brakkingsperiode vil være mest aktuell mot flerårig ugras. Helbrakk vil ha sterk effekt på nær sagt alle slike ugras. Figur 2.26 og 3.26 viser at man reduserer kveka betydelig ved gjentatt jordarbeiding gjennom vekstsesongen. Helbrakk er i dag lite aktuell, da må i så fall ugrassituasjonen være svært kritisk.
Ulike former for redusert jordarbeiding har blitt svært utbredt i mange land de siste tiårene. Primært er det ønske om bedre vannhusholdning og reduksjon av erosjon som ligger bak en slik praksis. Imidlertid har utstrakt bruk av redusert jordarbeiding i mange av disse områdene bidratt til en økning av Fusarium-angrep og innhold av mykotoksiner i korn. Det er vist også i norske forsøk at redusert jordarbeiding, da spesielt vårharving eller direktesåing kan bidra til økt angrep av Fusarium og andre sjukdommer. Dette viser betydningen av tradisjonell jordarbeiding/pløying for bekjemping av plantesjukdommer; ved å begrave infiserte planterester blir de nye plantene mindre utsatt for smitten. Likevel er det ikke generelt slik at jo mindre jordarbeiding desto mer sjukdom. Jordarbeidingspraksis vil kunne påvirke forekomsten av sjukdommer på flere måter. I tillegg til den direkte effekten på planterestene finnes det flere indirekte effekter av jordarbeiding.
Et av formålene med jordarbeiding er å løse opp pakket jord og legge til rette for god rotvekst. Bruk av tunge maskiner, særlig på våt jord, gjør at problemet med jordpakking flytter seg nedover i jorda. Det finnes eksempel på at bruk av tungt maskineri i en byggåker økte forekomsten av sjukdommer i hvete påfølgende år. Også to år etter, i bygg på samme sted, var forekomst av sjukdom forhøyet. I Frankrike er det rapportert om forhøyet forekomst av Rhizoctonia solani på bønner dyrket etter poteter som ble høstet med tungt maskineri. En annen type pakking er forbundet med det vi kaller plogsåle. Plogsåler kan favorisere flere ulike sjukdommer som f. eks. angrep av skarp øyeflekk. En dypere jordarbeiding, ned til 35-45 cm, kan bøte på problemer med plogsåler og fremme dypere rotvekst som igjen kan gjøre at man unngår tørkestress. I sin tur kan dette være med å forebygge sjukdomsproblemer. Dypere jordarbeiding er vist å kunne motvirke angrep av nematoder, men også soppsjukdommer.
Å begrave planterester gjennom pløying vil kunne hindre en del sjukdomssmitte i å spre seg videre til nye planter. Virkning av pløying til 25-30 cm dybde på fordeling og dekking av planterester med soppsmitte er likevel ikke alltid entydig. Effekter av ulik type jordarbeiding på sjukdommer i korn har vært nøye studert i England. Endring av jordarbeidingspraksis fra pløying til direktesåing har betydd lite for forekomst av rotdreper i bygg og hvete, men med variasjoner mellom år og steder. Dette forklares med at jordarbeiding påvirker mange andre faktorer. For eksempel vil jord som ikke vendes, kunne ha høyere grad av antagonister mot rotdreper i toppjordlaget, samt høyere forekomst av fosfor og lavere forekomst av nitrogen i øvre jordlag enn jord som vendes. På den annen side vil ikke-pløyd eller direktesådd jord kunne fremme utvikling av rotdreper i tilfeller der denne jordarbeidingspraksisen fører til mer pakket jord. Infiserte planterester i øvre jordlag, samt vekst av flerårige grasarter som kan være bærere av sjukdommen, kan også virke fremmende på utvikling av rotdreper.
For sjukdommer som hveteaksprikk vil pløying være mer entydig positivt, da det her er god sammenheng mellom mengden av infiserte planterester og sjukdomsutvikling. Samme effekt vil vi kunne se for andre sjukdommer som smitter direkte fra planterester i jorda. Mjøldogg derimot, kan av og til utvikle seg mer på planter i pløyd jord enn på direktesådd. Dette har sammenheng med redusert nitrogenmineralisering i ikke-pløyd jord som igjen gir tregere vekst og mer glissent bestand. Mjøldogg smitter som kjent via sporer i luft, og ikke via planterester på bakken slik som hveteaksprikk.
Også for andre sjukdommer og i ulike kulturer betyr pløying at planterester med smitte blir dekket av jord. Smittespredning blir dermed hindret, og man får redusert sjukdomsforekomst. Denne formen for kontroll er viktig for eksempel for Rhizoctonia solani, som kan forårsake sjukdom i flere ulike vekster (potet m. fl.) og salatbladskimmel.
En del overlevelsesorganer av soppsjukdommer bør likevel ikke «begraves» for dypt (under vanlig pløyedybde). Under dette nivået vil slike soppstrukturer (for eksempel sklerotier) bevares bedre enn i det mer aktive, øvre jordlaget. Det er bedre at inokulum/sjukdomssmitte blandes inn i det øvre jordlaget der det vil kunne spire i jorda eller brytes ned av andre mikroorganismer uten å nå vertsplantene.
Jordarbeiding mellom kulturene er også viktig for skadedyrkontroll. Skadedyr som er på planterestene når de pløyes ned, vil bli ødelagt slik at neste generasjonen reduseres. Eksempler på dette er mange bladlusarter og minerende flue- og møllarver som ikke kan rømme planten når denne pløyes ned. På samme måten er puppestadiet hos mange insekter å finne på plantedeler eller -rester. Også disse vil ta skade av jordarbeiding.
Skadedyr som insekter og snegler som tilbringer deler av utviklingen i jorda, vil ta skade av jordarbeiding. Disse har funnet fram til et jordsjikt eller annet bestemt sted i jorda hvor de vil ha best mulige overlevelsesvilkår. Ved pløying blir de både knust og forflyttet til andre sjikt/steder enn de selv har valgt. Dette vil som regel være ugunstige forhold for overlevelse på grunn av frysing, uttørking, lysforhold, m.m. Predasjon, ikke minst fra fugler, øker også ved pløying. Det er ikke tilfeldig at fuglene følger etter plogen, der mange godbiter kommer til syne!
Forsøk i England har vist at minst halvparten av kålfluene som overvintrer, ble drept ved vårpløying, mens ca. 80 % ble drept ved høstpløying. Det er viktig å huske på at jordarbeiding sprer nematoder i åkeren, og fra jorde til jorde med redskap. Dersom smitte er til stede, må all redskap som kommer i kontakt med jord, rengjøres omhyggelig før den flyttes.
For sjukdomsorganismer og skadedyr som lett beveger seg, uansett om de har få eller mange vertsplanter (se Figur 3.1 «C» og «D») er vekstskifte altså et lite effektivt tiltak. Flere svært problematiske skadegjørere for økologiske bønder, for eksempel tørråte på potet og kålflue på kålvekstene, hører hjemme her. Innenfor forebyggende ugraskontroll spiller derimot et optimalt utformet dyrkingssystem en svært viktig rolle.
Eksempler på aktuelle forebyggende tiltak som har effekt mot denne gruppen av skadegjørere, er en sterk (motstandsdyktig) kulturplante, størst mulig mangfold av planter på gården, og bruk av jorddekke. Når det gjelder kulturplanten, skal vi diskutere både valg av arter og sorter, og hvordan kulturplanten bør etableres på jordet (gjødsling og kalking, såmengder m.m.), slik at man i størst mulig grad unngår problemer med skadegjørere. For noen skadegjørere, og da tenker vi mest på ulike sjukdommer, vil også høstetid være en viktig faktor for å redusere avlingstap mest mulig. Under mangfold (diversitet) vil vi komme inn på bruk av arts- og sortsblandinger, dessuten bruk av underkultur og andre systemer med dekkekulturer.
I første kapitlet i denne boka var vi inne på et emne som ble kalt produksjonsøkologi (se tekstboks 1.7), og det ble bl.a. nevnt at valg av «rett» sort kan være avhengig av om man driver økologisk eller konvensjonelt. I ugrassammenheng vil optimal etablering av en mest mulig konkurransesterk kulturplante kunne bety mye. Konkurranseevna til kulturplanten, sammen med jordarbeiding, er selve fundamentet for ugraskontroll i et hvert driftssystem. Fordi man i økologisk landbruk ikke har så effektive direkte tiltak som ved konvensjonelt landbruk, blir dette fundamentet ekstra viktig. I ugraskampen handler konkurranseevne om to typer faktorer, plantens iboende egenskaper, og hvordan utenforstående faktorer påvirker planten.
For mange sjukdommer og skadedyr vil også ulike faktorer omkring kulturplanten være svært viktige. Ikke minst vil et optimalt sortsvalg være viktig, men vi vet også at ulike aspekt ved etableringa på jordet kan ha stor betydning.
Det vanskelige valget bonden må gjøre, omfatter et helt sett av kriterier hvor både skadegjørere, næringsforsyning m.m. inngår i en helhetlig vurdering. Det hjelper ikke om planten unngår sjukdommer, hvis den omkommer av konkurranse.
Valg av arter og sorter
Ugras
Valg av kulturplante påvirker i høy grad vekst og utvikling hos ugraset. Planter som vokser fort og skygger godt, hemmer ugraset langt bedre enn planter som vokser sent og/eller lager åpne, lyse bestander. Også sortsvalget kan være av stor betydning. Innen kornartene vil for eksempel kortstråete sorter oftest dekke dårligere enn sorter med lengre strå.
Generell gruppering av konkurranseevna til ulike kulturplanter:
Det kan være store forskjeller i konkurransekraft mellom de nevnte grupper av vekster. Det er for eksempel velkjent fra konvensjonell dyrking at havre og bygg konkurrerer mye sterkere med ugraset enn hva vårhvete gjør. Innenfor grønnsaksartene, for eksempel mellom gulrot og hodekål, er forskjellene enda større.
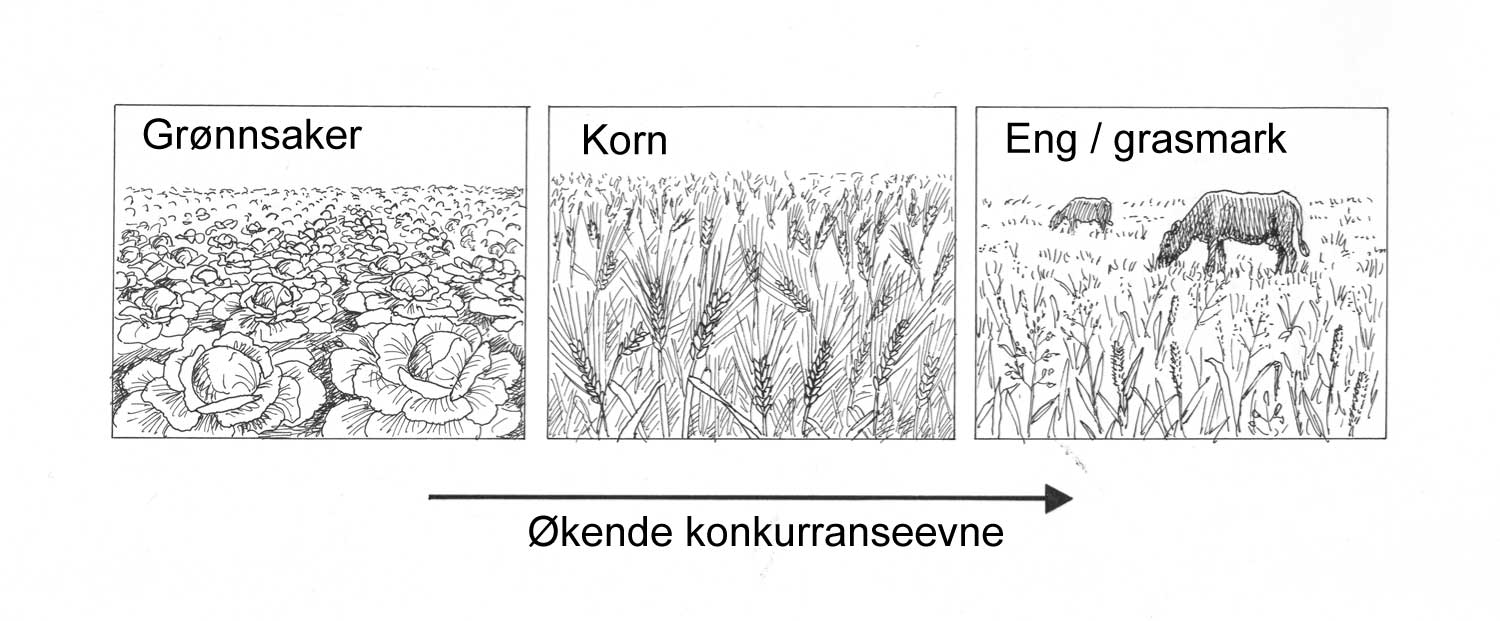
Ulike kulturveksters konkurranseevne vil variere med bl.a. sortsvalg, hvor plantene dyrkes og gjødselnivå. Et svensk forsøk har vist at ved sterk nitrogengjødsling konkurrerte bygg bedre mot ugraset enn hvete, mens det motsatte ble observert ved svak nitrogengjødsling. Høstkorn blir ofte vurdert som mer konkurransesterkt enn vårkorn, men denne vurderingen er primært gjort på ugras som spirer og etablerer seg om våren. På den annen side vet vi at vinterettårige og toårige ugras som spirer om høsten, kan skape store ugrasproblemer i høstkorn. Det er i kulturer som dekker jorda raskt, fortrinnvis korn og oljevekster, at konkurranseevna kan utnyttes som et betydelig bidrag i ugraskampen. Bruk av for eksempel en konkurransesterk kålsort kan også være viktig, men da er den største gevinsten at selektiviteten ved mekaniske og termiske tiltak forbedres. Størst mulig kulturplanter og små ugrasplanter er et godt utgangspunkt for ugraskampen.
Det er store forskjeller mellom sorter innen en og samme kulturvekst. En gjennomgående erfaring er nok at sorter som klarer seg bra ved konvensjonell dyrking, også klarer seg bra ved økologisk dyrking. I Norge har det ikke vært systematisk testing av ulike sorters konkurranseevne i økologisk dyrking. Sortsprøvinga for økologisk landbruk har hatt størst aktivitet innen potet og korn, og for begge disse vekstene har resistens mot sjukdommer vært prioritert når det gjelder plantevern. I Sverige er det publisert resultat av sortsprøvinger for økologisk dyrking av korn, potet og ert hvor også konkurranseevne er tatt med.
Hva er karakteristisk for en konkurransesterk sort? Som eksempel kan nevnes at en kornsort med hurtig bladvekst og sterk høydevekst vil konkurrere godt med ugraset.
Sjukdomsorganismer
Valg av kulturvekst og sort er ofte bestemmende for hvilke sjukdommer som opptrer i en åker eller et område. I mange tilfeller er sjukdomsorganismene artsspesifikke, dvs. de angriper bare en type kultur eller art. De vil ofte ikke kunne overleve særlig lenge på et skifte eller i en åker uten å ha en vertsplante å leve på. En del vil kanskje leve på døde planterester en sesong eller to, for så å dø ut. Lar man være å dyrke vertsplanten et par år, vil man på denne måten kunne sanere det infiserte plantematerialet. Dette er selvsagt en viktig effekt av vekstskifte og blir nærmere omtalt i andre avsnitt.
Valg av mest mulig resistent sort kan bety svært mye for forebygging av sjukdomsangrep. Det finnes eksempler på at selv om sterkere sorter er tilgjengelige, brukes sjukdomssvake sorter, begrunnet med at «markedet vil ha denne sorten». Den økte bruk av soppmidler som dette medfører, vekker til tider sterke reaksjoner i det samme markedet. For de fleste vil det imidlertid være mulig å velge sorter som er sterkest mulige mot de dominerende sjukdommene i området. I mange år har det pågått foredling i Trøndelag av sorter for å forbedre resistensen mot grå øyeflekk, som kan være svært destruktiv i dette området. Dette er den viktigste soppsjukdommen i bygg i Trøndelag. På Østlandet vil man gjerne velge en kornsort som er mest mulig resistent mot mjøldogg og byggbrunflekk. Resistensforedling er et langsiktig og kontinuerlig arbeid. En sort som er helt resistent mot en sjukdom, kan få sin resistens brutt ned. Særlig mjøldoggresistens i bygg og hvete har en tendens til å brytes etter en tid. Dette skyldes at også den resistente sorten blir utsatt for store mengder soppsporer og at disse sporene vil kunne ha en viss genetisk variasjon. De mest tilpassede vil kunne overleve. De sporene som har fått endret sine genetiske egenskaper slik at de greier å leve på planten, har altså brutt plantens resistens, og vil dermed oppformeres i store mengder og spre seg videre. Et konvensjonelt dyrkingssystem der man dyrker samme sort over store arealer vil være særlig utsatt for å få brutt resistensen. Det vil ofte være slik at planter som har en viss grad av resistens, basert på flere gener eller egenskaper, vil beholde resistensen i langt sterkere grad enn i det ovennevnte tilfellet. Fullstendig resistens er ofte basert på det som i forrige kapittel ble kalt spesifikk resistens og baserer seg på et eller få gener, noe som altså har sine svake sider. Feltresistens, eller resistens som baserer seg på flere gener, vil være å foretrekke, særlig dersom samme sort dyrkes hyppig over større arealer.
Skadedyr
Plantenes resistens mot skadedyrangrep kan arte seg på tre forskjellige måter. Vi kaller disse for antixenose, antibiose og toleranse. «Antixenose» er når et skadedyr som vanligvis angriper et planteslag, ikke angriper den resistente sorten. Det kan for eksempel være på grunn av små forskjeller i plantens utseende, lukt eller smak som gjør at skadedyret ikke «liker» planten.
Vi bruker uttrykket «antibiose» når skadedyret angriper planten som vanlig, men dyret tar skade av planten. Planten kan f.eks. ha en kjemisk sammensetning som er til skade for dyret eller den kan ha en fysisk utforming med kroker eller klebrige stoffer som skader eller fanger dyret.
«Toleranse» er mindre dramatisk enn antibiose. Med toleranse menes at planten får like mye angrep som andre sorter, og dyret klarer seg like bra, men planten overlever og produserer en god avling til tross for angrepet.
Selv om det arbeides mye med å utvikle plantesorter som er resistente mot skadedyr er det få eksempler på bruk av resistens i norsk landbruk. I utlandet er det utviklet resistens mot trips i kålvekster og delvis resistens mot kålfluer. Det finnes også sorter av salat som er resistente mot bladlus. I Norge har vi en potetsort som har toleranse mot sikadeangrep, men sorten er mindre populær hos norske forbrukere. Dette er et vanlig problem i forbindelse med utvikling av resistente sorter. Det kan ta mange år i et foredlingsprogram å få til en sort med resistens, men jobben er ikke ferdig før sorten også har fått den rette matkvalitet, farge og konsistens som forbrukerne vil ha.
Mot nematoder er resistens ofte meget viktig. Resistens kan fungere bedre enn karens for å rense et jorde for potetcystenematoder. Dette skal vi ta i mer detalj i Bind 2 om potet.
Rent og friskt frø og formeringsmateriale
Ugras
En del land har egne lover for å hindre spredning av ugras. Norge har ingen generell ugraslov, men vi har to forskrifter (Forskrift om floghavre, av 25.03.1988 og Forskrift om såvarer, av 13.09.1999, begge med endringer og hjemlet i Lov om matproduksjon og mattrygghet, av 19.12.2003) som skal redusere spredningen av ugras, blant annet ved at innholdet av ugrasfrø i såvarene vi kjøper, ikke må overstige visse definerte verdier.
Et aspekt i ugraskampen er altså å unngå «blindpassasjerer» i frøposen, et annet er å bruke frø med optimal spireprosent og spirehastighet. Forsøk i utlandet, blant annet danske forsøk i hvete og bygg, har vist at kvaliteten til såfrøet er av stor betydning for konkurranseevna mot ugras. Såfrø solgt i Norge, unntatt såkorn, testes for spirehastighet, men du finner som regel ikke opplysninger om resultatet av disse testene på frøposene. Det er rimelig å anta at kvaliteten på såfrøet er enda viktigere ved økologisk enn ved konvensjonel dyrking fordi man der ikke kan «kompensere» ved å sprøyte mot ugraset.
Sjukdommer
Sjukdommer kan spres videre på ulike måter. Noen sjukdomsorganismer følger med utgangsmaterialet vi har når vi sår eller planter. Det kan altså være fort gjort å så sjukdomsorganismen ut sammen med såkornet, eller sette den ut med settepoteten. Bruk av såkorn, såfrø, småplanter og settepoteter som er kontrollert og testet for sjukdomssmitte kan være en god investering med tanke på senere angrep av sjukdom.
Det finnes også regler for økologisk produksjon av såvarer/utgangsmateriale. Utgangsmaterialer skal være produsert økologisk. En del sjukdomsorganismer har såkornet som eneste smittevei, mens en del andre overlever i tillegg i planterester på åkeren og kan spre seg videre fra disse. Muligheter for kjemisk beising har ført til lite foredlingsarbeide når det gjelder resistens mot sjukdommene i førstnevnte kategori. Resultater fra resistenstesting mot for eksempel stinksot i høsthvete og stripesyke i bygg viser imidlertid at det finnes forskjeller i resistens mot disse sjukdommene. Konkrete tiltak med hensyn på produksjon av korn og såkorn vil bli nærmere omtalt i Bind 3.
Skadedyr
Når man kjøper eller avler opp planter, er det meget viktig at plantene inspiseres av en som kjenner til skadedyrene før plantene tas i bruk, for å unngå at en tar med seg smitte ut i det nyutplantede feltet. Bruk av egne settepoteter kan også være en mulig smittekilde og bør ikke skje ukritisk.
Kulturplantens plassering på gården
Ugras
«Fysisk» plassering av en kulturvekst på gården har begrenset betydning for ugraset. Den effekten man eventuelt kan se på ugraset, er av mer indirekte art, ved at kulturplanter som etableres på en mindre heldig plass på gården, vokser dårligere og dermed stiller svakere i konkurransen med ugraset.
Sjukdommer
En gårdbruker som kjenner gården og jorda si godt, vil ha et godt erfaringsgrunnlag for valg av vekster på de ulike skiftene på gården. Dette valget vil ofte tas med utgangspunkt i jordas egenskaper og kulturens vekstkrav. I visse tilfeller kan plasseringen få betydning for sjukdomsangrep. Velger man for eksempel å dyrke samme sort og vekst som naboen, kan det være en fordel å plassere denne veksten et godt stykke unna naboens åker, slik at man får en fysisk barriere som eventuelle sjukdomsorganismer må krysse. For sjukdomsorganismer som lett spres med vind, vil ikke dette være særlig effektivt, men for sjukdomsorganismer som spres hovedsakelig med regn og/eller med vind bare over korte avstander, vil det kunne bety en del. Tidligpoteter bør aldri dyrkes ved siden av senere poteter. Tidligpotetene rekker ikke å få så mye skade av tørråte, men kan selvsagt være infisert og fungere som en kraftig kilde til videre spredning av soppen over til de sene potetene der den vil få tid til å gjøre større skade. På samme måte bør høst- og vårhvete heller ikke ligge inntil hverandre fordi sjukdomsorganismer vil opptre tidligere i de høstsådde feltene, og bidra til raskere infeksjon av de vårsådde feltene. Bruker man slik kunnskap til å forsinke et angrep mest mulig, vil plantene være sterkere når angrepet først kommer. Vind kan som sagt spre sjukdommer, men kan også være med på å dempe utvikling av enkelte sjukdommer. Soppsporer trives ofte i fuktige bestander, og plassering av for eksempel potet på skifter som er mer utsatt for vind, vil kunne gjøre at bestanden tørker raskere opp. For jordbær er det også positivt med rask opptørking for å dempe angrep av gråskimmel. Vi kan bedre opptørking ved å legge planteradene langs med gjeldende vindretning. Dette gjøres av og til også i frukthager (hvis det er praktisk mulig) for å få hurtigere opptørking og redusere perioden med fuktige forhold.
Kjennskap til forskjeller i jordas pH kan være en faktor som bør spille inn ved valg av skifte for dyrking av for eksempel kålvekster. Jord med høy pH vil kunne virke forebyggende mot angrep av klumprot. Motsatt vil jord med lav pH favorisere denne soppen. På lengre sikt vil man selvsagt kunne bruke kalking som et forebyggende middel, men for plassering av vekster et aktuelt år, vil slike forskjeller ha betydning. Den generelle mikrofloraen i jord vil kunne påvirke de jordboene sjukdomsorganismene. I mange sammenhenger er det funnet at visse åkre har en naturlig sjukdomshemmende effekt. Dette er et spennende fenomen, men det er vanskelig å kartlegge og handle ut fra dette. Generelt vil en rikere mikroflora og -fauna i jord være sterkere mot jordboende sjukdomsorganismer som for eksempel Fusarium-arter som angriper en rekke planter, bl.a. korn og potet. Når dette er sagt, vil man raskt komme over på tiltak som endrer mikroflora i jord, som gjødsling, drenering, jordarbeiding. Effekter av slike tiltak vil vi komme tilbake til.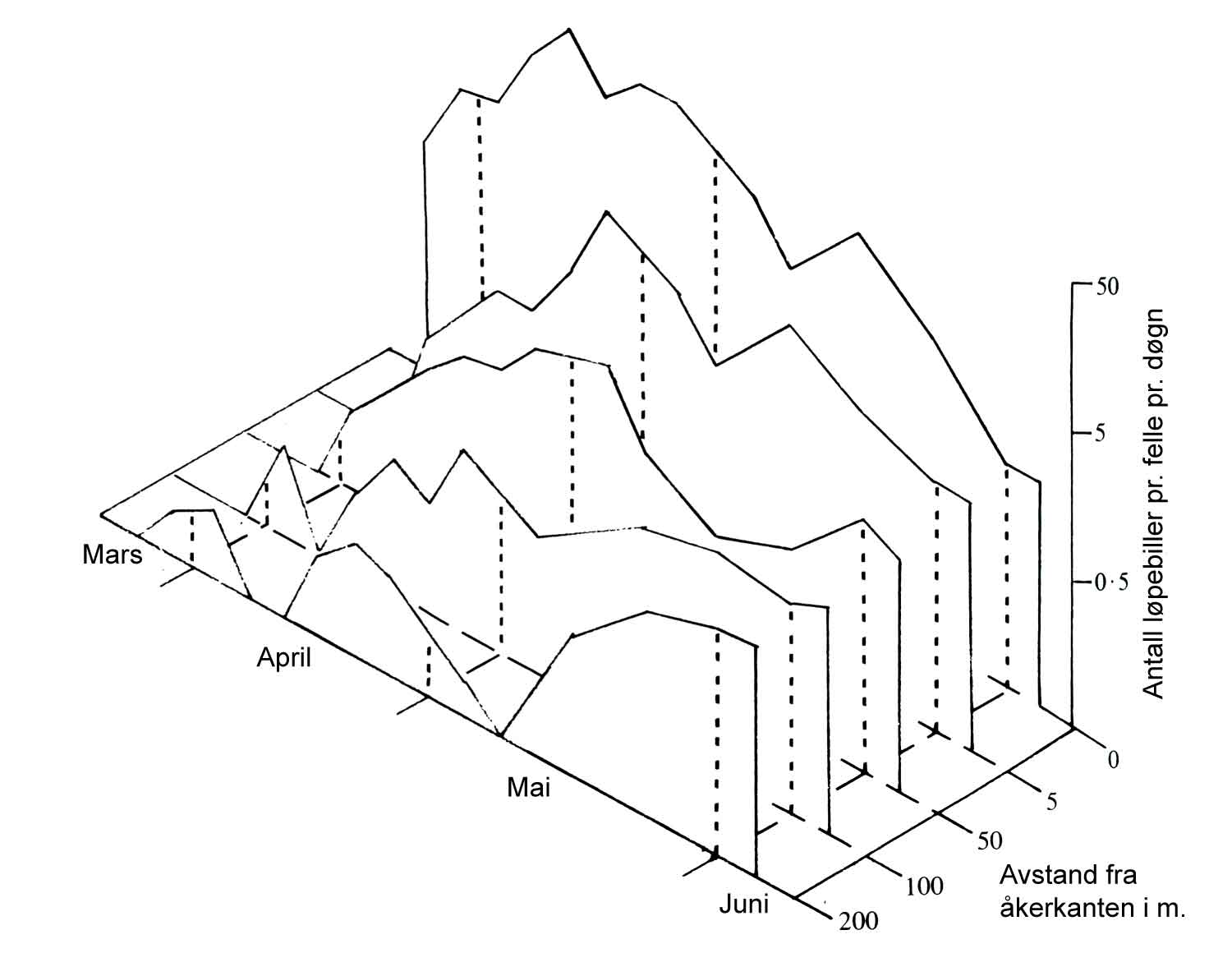
Skadedyr
For flerårige kulturer som frukt og bær, og delvis også for andre kulturer vil agronomiske hensyn veie mer enn risiko for skadedyr for hvor den enkelte kulturen plasseres på gården. I den grad skadedyrsituasjonen kan påvirke valget, er det flere ting å ta hensyn til. Hva som er best i hver enkelt kultur, vil du finne mer om i kulturheftene, men eksempler på hva man skal vurdere er jordtype, nærhet til kantvegetasjon, nærhet til beslektede kulturer og vindretning. Et eksempel på dette er bladlus i korn, som er mer utsatt for naturlige fiender når det er kort avstand fra kornåkeren til åkerkanten hvor nyttedyrene, f.eks. løpebiller, kommer fra (figur 3.29).
Bladlusa blir ført passivt med vind, og på grunn av turbulens lander den ofte lenger fra f.eks. skogkanten. Situasjonen er omvendt for gulrotfluen. Den har tilholdssted i plantene i åkerkanten, og i tillegg er den en svak flyver. Derfor får vi mest angrep på gulrot i kantradene. Da er det om å gjøre å plassere gulrotåkeren så langt fra kantvegetasjonen som mulig (figur 3.30).
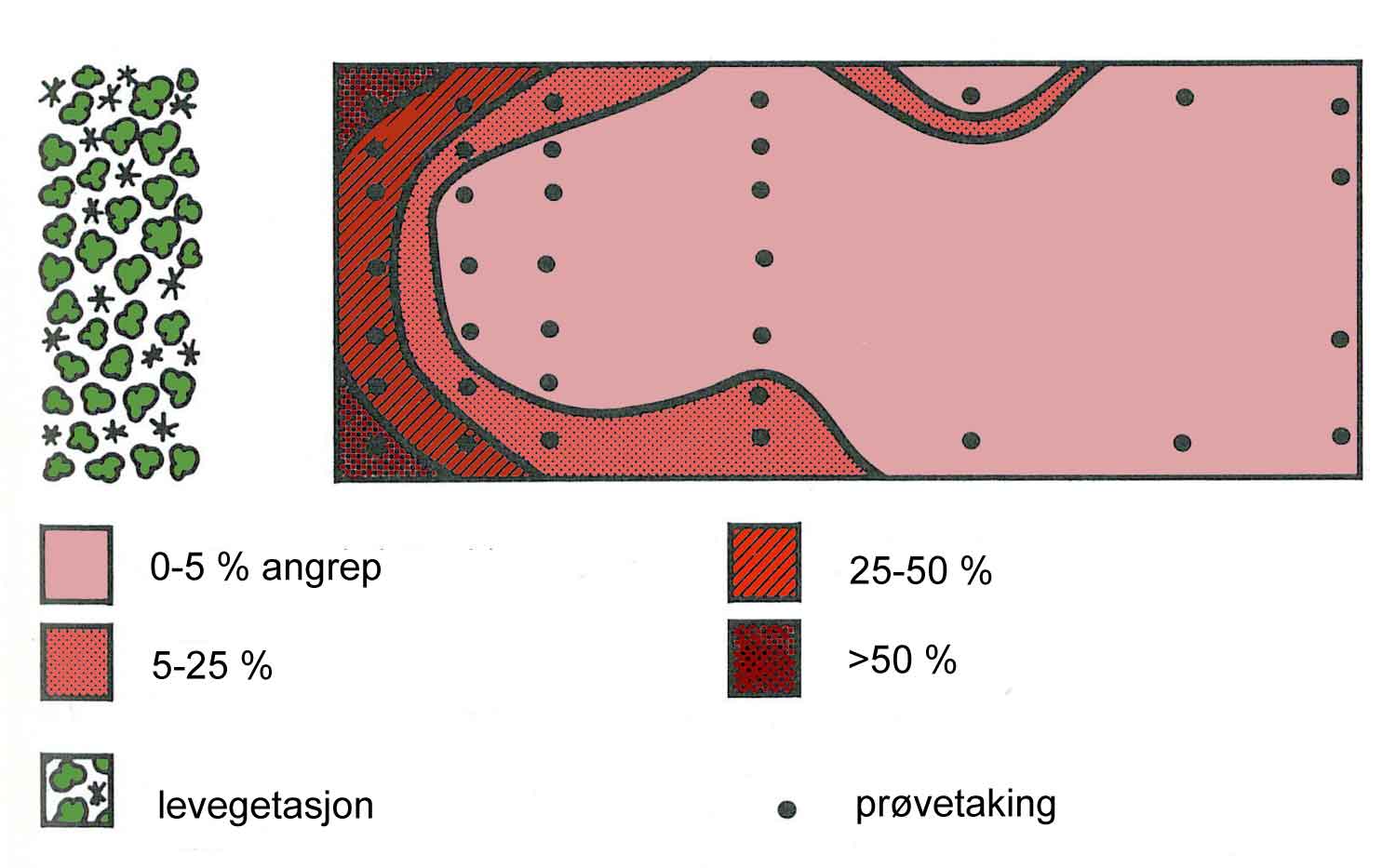
Ugras og gjødsling
Gjødsling virker inn på de ulike plantenes vekst- og konkurranseevne. Ugraset reiser som kjent med næring fra jorda. Følgelig blir det mindre tilbake til kulturplantene som av den grunn vokser dårligere. Det ligger da nært å tro at tilførsel av mer gjødsel kan motvirke dette ved at kulturplantene får den næringen de trenger og derved utkonkurrerer ugraset. At dette også kan slå til i praksis, for eksempel når ugrasfloraen er sammensatt av arter som er lite i stand til å utnytte større næringsmengder, er sikkert nok, men resultatet av økt gjødsling kan også bli det motsatte, altså at kulturplantene blir undertrykket av ugraset. Dette er et felt vi vet for lite om i dag, men mange av dagens ugras utnytter for eksempel nitrogengjødsling like godt eller bedre enn kulturplantene (se tekstboks 3.9). Dette er en konsekvens av at ugrasfloraen gradvis har tilpasset seg dagens moderne landbruk. I svenske forsøk har det vist seg at flere ettårige ugras, for eksempel meldestokk, og ugraspopulasjoner i sin helhet, reagerer mer positivt på nitrogengjødsling enn kulturvekstene, som i dette tilfelle var bygg, havre og hvete. Tabell 3.10 gir en oversikt over «taperne», dvs. arter som tidligere var vanlige ugras, men som det nå har blitt mindre av bl.a. pga. økt gjødsling eller bedre drenering. Andre faktorer enn gjødsling har nok også vært medvirkende årsak til at de nevnte artene har gått tilbake. I økologisk landbruk gjødsles det mindre enn i konvensjonelt landbruk, og man kan forvente seg at ugrasfloraen vil tilpasse seg dette ved at en del mer sjeldne arter dukker opp igjen.
Mange undersøkelser har vist at det er vanskelig «å gjødsle bort ugrasproblemet». Tabell 3.11 presenterer et forsøk som ble utført i kar med ulike sammensetninger av havre og ugras (åkersennep). Ugrasmengda i kara med havre ble variert fra 0 ugrasplanter til enten 5 eller 10 ugrasplanter. Hele forsøket ble i tillegg kombinert med 3 ulike gjødselnivåer, lite (G1), middels (G2) og mye (G3) gjødsel. Hvis havren stod alene, medførte økt gjødsling ganske klart økt opptak av nitrogen i havreplantene og økte avlingstall. Når havren stod sammen med 5 eller 10 ugrasplanter, økte mengda av opptatt nitrogen, og enda tydeligere avlinga, lite eller ikke ved sterkere gjødsling. Ugraset gjorde på den annen side seg helt klart nytte av den økte gjødslinga.
Tabell 3.11 Hvordan økende gjødselmengde (G1 minst, G3 mest) påvirker konkurranseforholdet mellom havre og ulike tettheter (0, 5 eller 10 planter per kar) av ugras (åkersennep) (etter Alkämper 1976)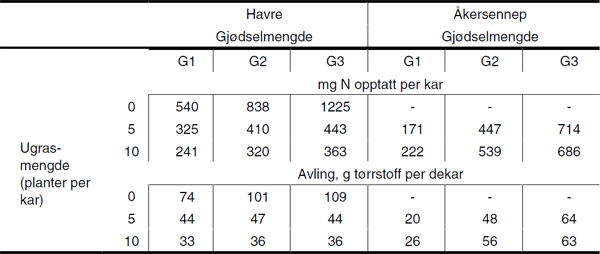
Forsøk utført på friland har også vist at det er ugraset som ofte tar opp den økte gjødselmengda. I et forsøk i Sverige ble det på 2 lokaliteter gjødslet med enten 4,5 eller 9,0 kg nitrogen per dekar. Halvparten av forsøksrutene ble luket, mens ugraset ble stående på den andre halvdelen. Figur 3.31 viser, analogt til potteforsøket over, at hvis det var mye ugras i feltet, gjorde ugraset seg mer nytte av den økte gjødselmengda enn kornet.
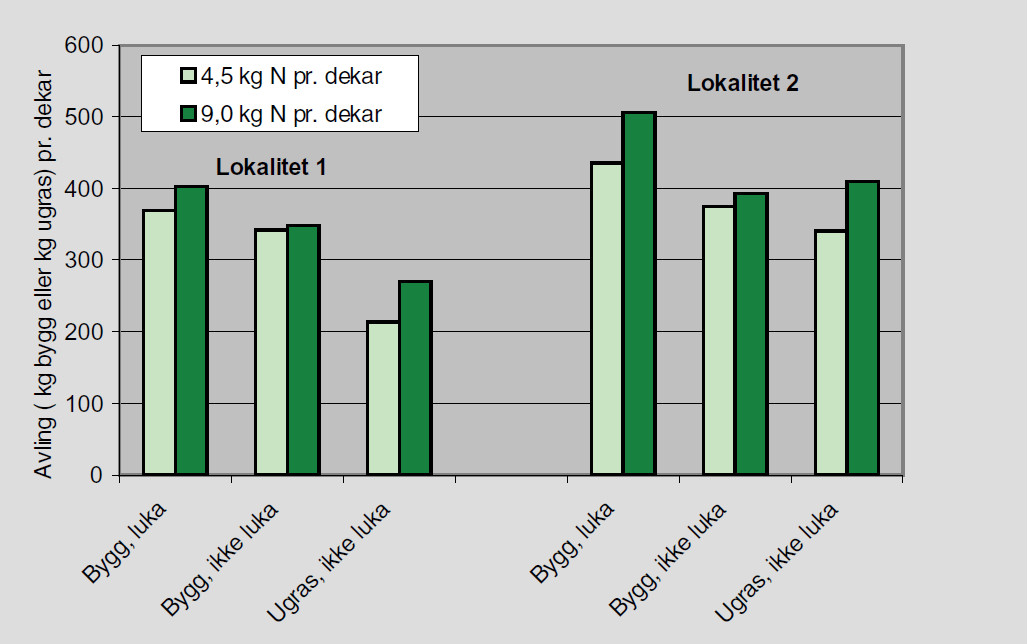
Tabell 3.10 Ugrasarter som det har blitt mindre av (noen nesten
forsvunnet) pga. sterkere gjødsling eller bedre drenering (etter Håkansson 1995)
| Art | Årsak til tilbakegang | |
| Økt gjødsling | Økt drenering | |
| Storengkall | x | - |
| Åkermarimjelle | x | - |
| Lodnefaks | x | - |
| Sanddodre | x | - |
| Sandskrinneblom | x | - |
| Engsmelle | x | - |
| Linbendel | x | - |
| Åkersnelle | x | - |
| Gåsemure | - | x |
| Åkersvinerot | - | x |
| Vasslirekne | - | x |
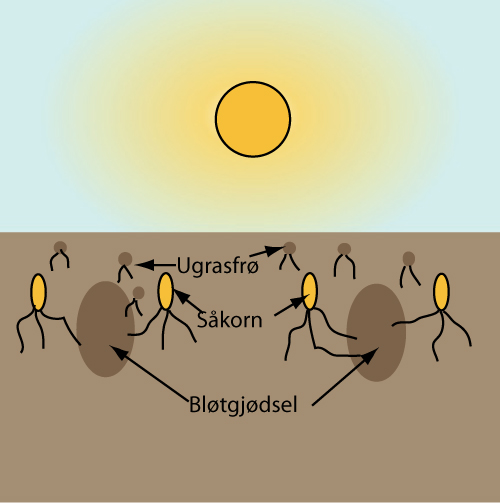
Ugras og grøfting
Grøfting av vassjuk jord er et viktig tiltak for å bedre kulturplantenes konkurranseevne i forhold til ugraset. Mange ugras trives særlig godt på våt jord, f.eks. hønsegras, vassarve, krypsoleie, åkersvinerot og sølvbunke. Den slags ugras blir ikke borte fordi om jorda blir grøfta, men de mister mye av fordelen de har av å vokse godt på vassjuk jord ved at kulturplantene generelt vokser best på godt drenert jord og dermed blir mer konkurransesterke. Dessuten får vi bedre vilkår for å arbeide jorda skikkelig og til rett tid, både vår og høst, og vi får bedre effekt av hakking og radrensing i selve vekstperioden. Alt dette gjør at ugras som i og for seg vokser bra på tørrere jord, blir mindre brysomme når jorda er godt grøfta.
Ugras og kalking
Kalking virker i flere retninger på ugrasfloraen isolert sett. En del ugras kommer avgjort best til sin rett på sur jord, for eksempel linbendel, stemorsblomst, småsyre og engmose. Kalking vil derfor motvirke disse artene. På den andre siden vil heving av pH til et nivå som er rimelig for de fleste kulturplanter også være en fordel for mange ugrasplanter. Når kalking av sur jord likevel er en fordel totalt sett, henger det først og fremst sammen med at jorda får en bedre struktur og derved blir lettere å ha med å gjøre, og at kulturvekstene selv blir sterkere konkurrenter til ugraset.
Ugras og jordstruktur
Praktiske erfaringer tilsier at ugrasproblemene tiltar på strukturskadet jord. Dette skyldes nok ikke at ugraset trives bedre på dårlig enn god jordstruktur, men at det tåler slike vekstforhold bedre enn de fleste kulturplanter.
Type og mengde gjødsel skiller økologisk landbruk fra konvensjonelt landbruk. Grøfting og kalking har også stor innvirkning på ugrasfloraen, men dette er ikke faktorer som er spesifikt forskjellig for økologisk og konvensjonell dyrking. Optimale forhold for kulturplantene, slik at spesielt rotsystemet er godt utviklet, er kanskje viktigere for økologisk enn for konvensjonell dyrking pga. at plantene generelt har mindre tilgang til næringsstoffer.
Sjukdom og gjødsling
Næringstoffer kan påvirke forholdet mellom kulturplanter og plantesjukdommer på mange forskjellige måter. I den grad bonden kan kontrollere tilførsel av ulike næringsstoffer, kan dette også til en viss grad påvirke angrep og utvikling av sjukdom i vekstene.
Vekst og vitalitet påvirkes av plantenes næringstilgang, og dette kan videre påvirke mikroklimaet og plantens evne til å motstå eller å «komme seg» etter et sjukdomsangrep. Veksthastigheten kan påvirkes og gjøre det mulig å forkorte de mest mottakelige stadiene (for eksempel frøplantestadiet) eller regulere andre stadier (blomstring), samt påvirke modningsstadiene. Tilstrekkelig næringtilførsel gjør det lettere for plantene å utnytte et overskudd av vann i rotsonen.
Noen næringsstoffer har direkte toksisk effekt på patogener, som for eksempel urea.
Nitrogen
Høye nitrogen-doser kan gi planter som er mer mottakelig for angrep av biotrofe sopper (sopper som bare vokser på levende planter) enn lavere nitrogen-doser (f.eks mjøldogg og rustsopper i korn). Dette skyldes både lavere resistens som følge av plantas sterke vekst, og samtidig frodigere og fuktigere bestand som igjen er gunstig for soppen. Motsatt kan man ofte observere et noe lavere angrep av slike sjukdommer i systemer/tilfeller der nitrogentilgangen er noe mer begrenset. I tillegg til langsommere vekst av selve plantene vil også redusert nitrogentilgang gi tynnere og mer luftig plantebestand og mindre optimale forhold for sopputvikling. På den annen side vil en del fakultative parasitter (sjukdomsorganismer som også kan være saprofytter på dødt organisk materiale) kunne angripe sterkere på mindre vitale planter eller planter i svakere vekst, og altså profittere på lavere næringstilgang.
Det finnes utallige studier som viser effekt av ulike doser og tidspunkter for kunstgjødsel og virkning på patogener. Især manipulering med nitrogentilførsel i korn er nøye studert. Selv om det ikke er de samme muligheter for manipulering med spesifikk næringstilførsel i økologisk landbruk, kan det likevel være viktig å vite noe om næringsstoffer og virkning på plantesjukdommer. Tidspunkt for nitrogentilførsel har også betydning. Man bør unngå tilførsel av for mye tilgjengelig nitrogen sent i sesongen ved plantenes modningsstadium fordi det fører til utsatt modning og økt grønnmasse. Dette vil igjen forlenge tida med muligheter for skade av patogener og øke angrep og utvikling av sjukdom. I korn påvirker sen nitrogentilførsel spesielt hveteaksprikk og rust, men også mjøldogg kan øke under slike forhold. For epleskurv er det vist at sen sprøyting med urea (før bladfall) kan øke nedbrytningen av bladene, noe som igjen kan minske sporeproduksjonen av epleskurv.
Fosfor
Fosfor (P) har naturlig nok også effekt på plantesjukdommer siden dette næringsstoffet også er svært viktig i plantens metabolisme. Likevel er virkningen av dette næringsstoffet på plantens resistensmekanismer mer variable. Noen undersøkelser viser en direkte virkning av fosfater på plantesjukdommer. I en langtidsstudie i England (Rothamsted) fant man at gjentatt tilførsel av fosfatgjødsel både forsinket og minsket angrep av rotdreper i bygg. I USA har man vist at sterk tilførsel av fosfat har redusert forekomst av flatskurv i potet. Hveteaksprikk derimot, viste seg å øke etter økende tilførsel av fosfatgjødsel. Effekt av fosfor på sjukdommer ser ut til å være merkbar bare der det har vært underskudd på fosfor i utgangspunktet, og er altså et bidrag til økt plantehelse der denne har vært svekket av ubalansert næring. I noen tilfeller kan også modning fremmes med fosfattilførsel og dermed gjøre at veksten unnslipper sjukdomsangrep. Det finnes lite praktisk anvendelig informasjon om fosfor og plantesjukdommer. Tilgjengelige gjødselslag, plantens behov og jordas status blir derfor avgjørende for tilførselen.
Kalium
Effekten av kalium (K) avhenger i stor grad av næringsstatus i jorda, mengde kalium i tilført gjødsel og av den balanserende effekten fra andre næringsstoffer – spesielt nitrogen. Effektene på planten og videre på angrep av sjukdomsorganismene kan være både direkte og indirekte. Indirekte effekter kan være sårheling, økt frostresistens (også økt resistens mot snømugg), samt utsatt aldring. Fusarium-rotråte i luserne er blitt redusert ved økt tilførsel av kalium. Det samme er mjøldogg i bygg og hvete, samt bladskimmel i blomkål. Kaliumtilførsel har også vist seg å kunne redusere visnesyke i erter.
Organiske gjødselslag
Næringstilførsel av mer langsomtvirkende slag som for eksempel husdyrgjødsel, grønngjødsel osv. er oftest studert i forbindelse med jordboende patogener. Ved tilførsel av organiske gjødselslag, vil effektene være mer komplekse og påvirkes av effektene på jordstruktur etc. Noe av det som er nevnt for nitrogen, fosfor og kalium vil også kunne forklare effekter av organiske gjødselslag på plantesjukdommer, men virkningene vil være mer komplekse og vanskeligere å tilbakeføre direkte til et enkelt næringsstoff. Påvirkningen på mikrolivet i jorda vil være større, og flere indirekte effekter vil kunne oppnås gjennom dette. Store variasjoner etter jordtype og type av tilført materiale vil også forekomme. Humusstoffer er resultater av nedbrytningsprosessene av organisk materiale i jord ved hjelp av mikroorganismer. Nedbrytningen er i sin tur avhengig av tilgjengelighet av karbon og nitrogen, og prosessene bremses når tilgjengeligheten på disse blir lavere. Det finnes svært mange studier som viser effekter av humusstoffer på planter. Stoffene kan virke på næringsopptaket og virke som vekstregulatorer, øke frøspiring samt øke aktiviteten til mikrofloraen.
Humusstoffer kan altså virke på plantehelse gjennom å påvirke røtter og organismer i rotsonen. En rekke undersøkelser med tilførsel av humusstoffer til ulike jordtyper viser at effektene kan variere. I noen tilfeller ble planteveksten fremmet, og sjukdomsangrep av jordboende patogener som for eksempel Fusarium redusert, mens man i andre tilfeller fikk veksthemming av plantene. Resultatene var avhengig av jordtype. De varierende resultatene indikerer at tilførsel av næringstoffer gjennom organisk materiale vil være til dels uforutsigbart med tanke på virkning på sjukdomsframkallende organismer.
Likevel vil det svært ofte være gunstige effekter på mikrolivet generelt og på jordas bufferevne ved tilførsel av organiske gjødselslag. Forbedret jordstruktur påvirker rotvekst og plantens evne til opptak av næringsstoffer og vann, og dermed også plantens motstandskraft mot stress og sjukdom.
Sjukdom og kalking
Heving av jordas pH gjennom kalking kan påvirke plantesjukdommer direkte og indirekte. Klumprot er nok det mest kjente eksemplet med kalking som tiltak. Endringer i pH vil også endre tilgjengeligheten til andre næringsstoffer. Mikronæringsstoffene blir ofte mindre tilgjengelige ved økende pH.
Sjukdom og drenering
God drenering kan bety mye for mikroklimaet i plantebestandet, også i tilfeller der plantene ikke er direkte påvirket av for mye vann i jorda. Jordtyper med ulik dreneringsgrad vil ofte være ulike med hensyn på forekomst av sjukdom, også ved ellers like klimatiske forhold. En lett eller godt drenert jordart vil gi raskere opptørking og tørrere mikroklima, noe som igjen kan være med på å begrense sjukdomsutviklingen. God drenering på tyngre jordarter bør derfor også være med som en del av det forebyggende arbeidet mot sjukdom.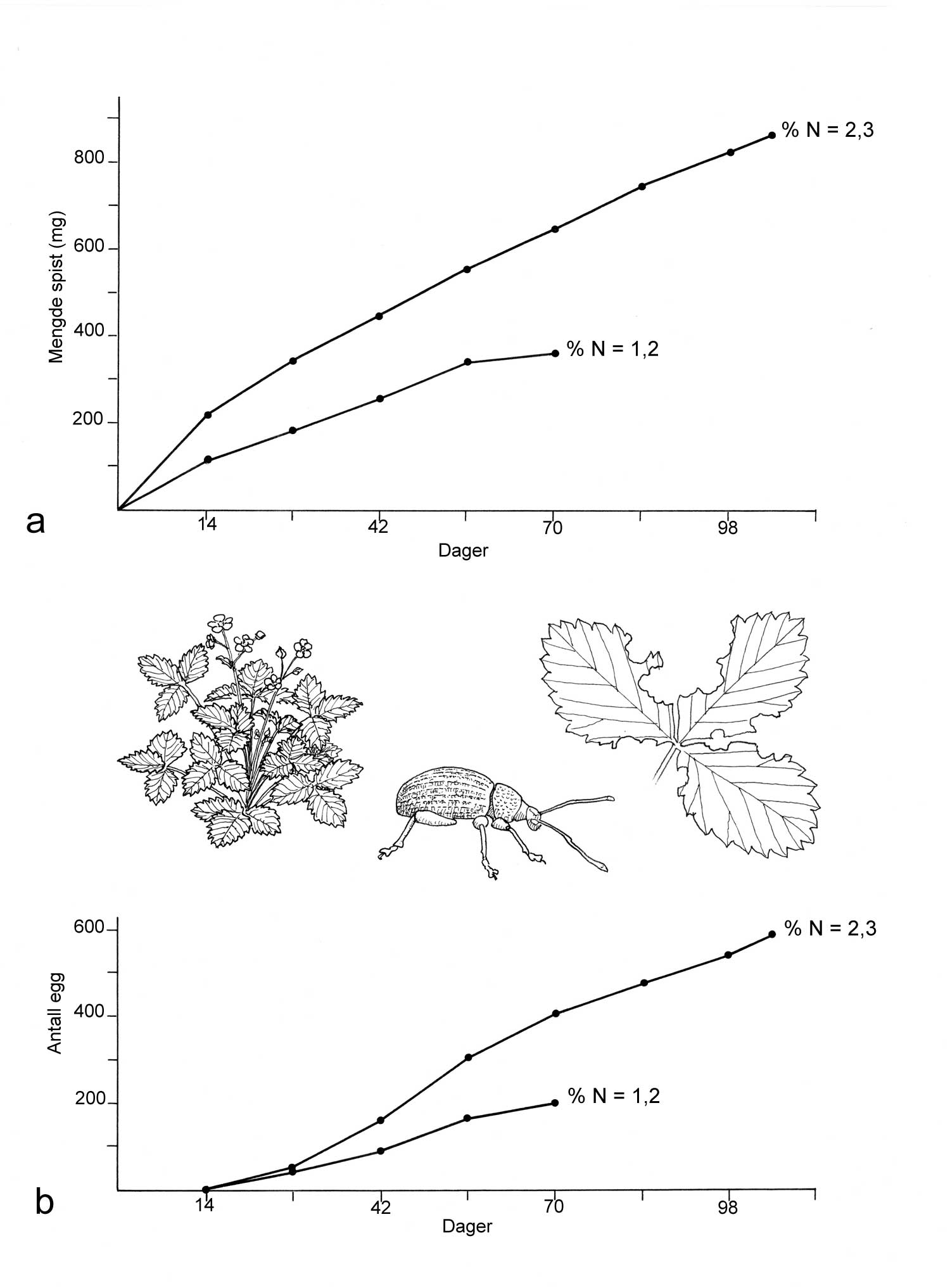
Skadedyr og gjødsling
Når nitrogeninnholdet i planten økes, forbedres vekst, reproduksjon og levetida hos insekter som lever av planten. Forsøk i Norge med rotsnutebiller i jordbær viste økt egglegging med økt nitrogeninnhold i bladene.
I tilsvarende studier i England med en bladtege ble veksten per matenhet spist, fordoblet når prosent nitrogen i maten ble fordoblet. Bladlus som har tvungent vertsskifte, forlater vinterverten når nitrogeninnholdet synker (figur 3.34). Mye tyder på at enkelte plantearter etter hvert har utviklet et lavt nitrogeninnhold som en form for forsvar mot insektangrep.
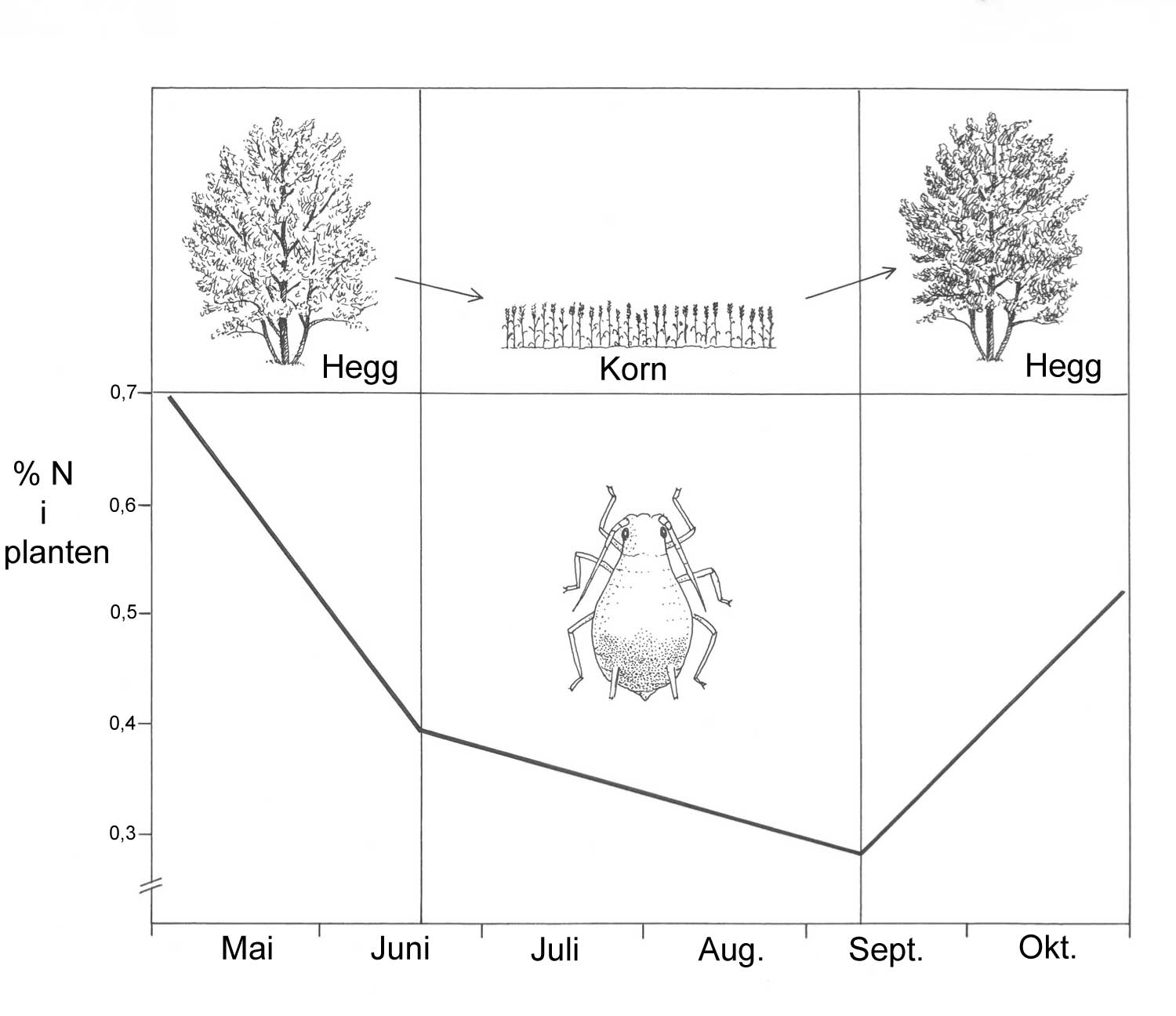
Ugras, så- eller plantetidspunkt
Gitt at vekstsesongen er lang nok, vil det for flere kulturvekster være bra for ugrassituasjonen å utsette tidspunkt for såing eller planting. Det er flere årsaker til at framveksten av ugras kan bli mindre på denne måten:
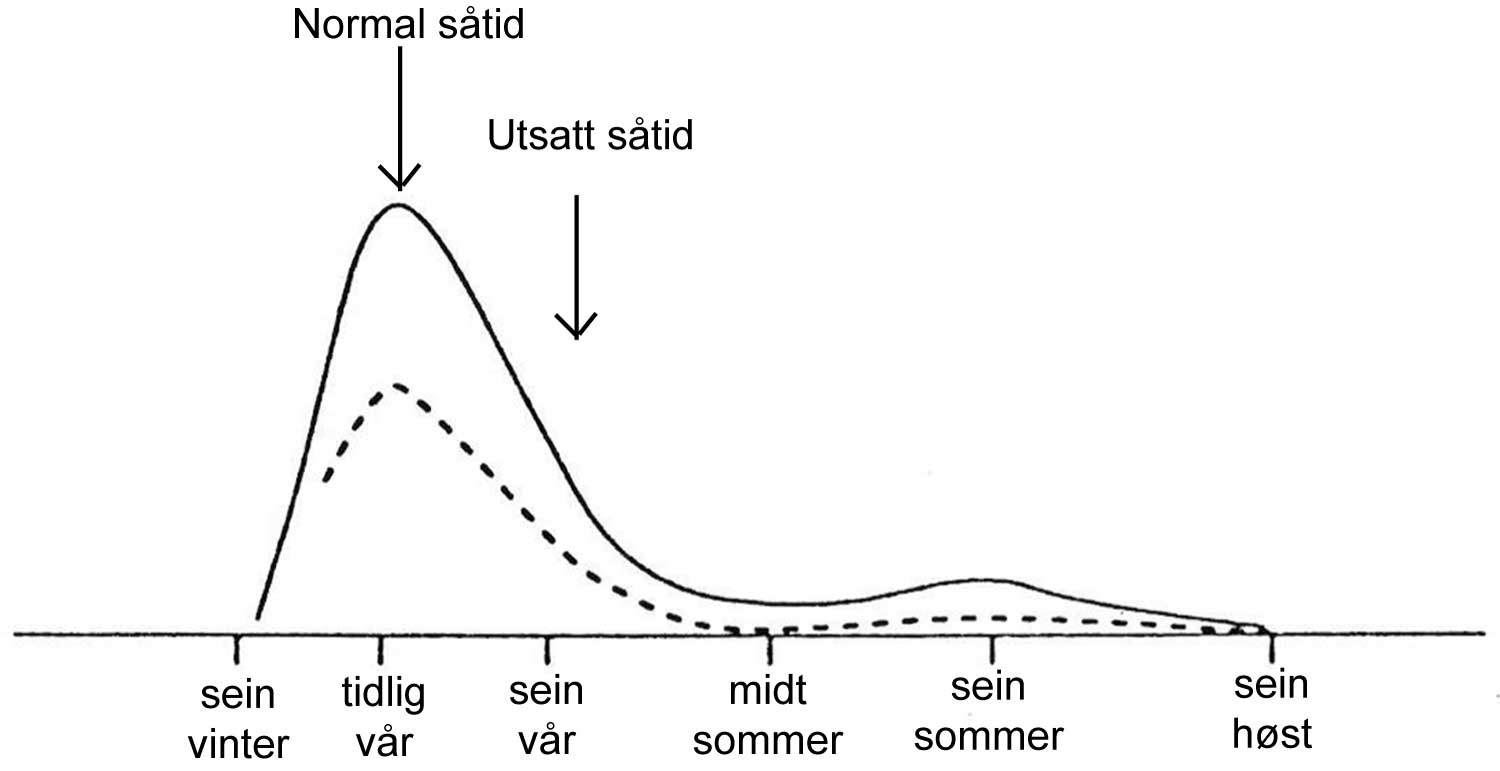
Å lage til såbedet om våren tidligere enn «nødvendig» for så å drepe ugraset som kommer opp ved å harve, kalles ofte falskt såbed (tekstboks 3.10). Hvis man har utstyr for det, bør man bruke flamming ved siste behandling rett før såing eller planting, for på den måten å hindre at mye nytt frø blir stimulert til å spire.
Prinsippet «falskt såbed» bygger på at man gjør såbedet klart for eksempel 10-14 dager før planlagt såing eller planting. Hvis man tar seg god tid, kan man etter å ha lagd til såbedet, ugrasharve, for eksempel en uke etterpå. Etter ytterligere en uke kan man svi ned ugraset med propan, for deretter å så eller plante kulturveksten. Det er viktig å være klar over at visse ugras, f.eks. tunbalderbrå, gjetertaske og åkerstemorsblomst, er vanskeligere å flamme bort enn andre (se kapitlet Direkte tiltak). Det vil alltid være en fordel å flamme på tørre planter og på tørr jordoverflate. Harving istedenfor nedsviing rett før såing eller planting er ikke så bra, fordi harvinga i større grad vil stimulere nye ugrasfrø til å spire. Imidlertid er det få som har flammeutstyr, men mange kan ha nytte av falskt såbed oppnådd med harving.
I et avsluttet norsk forskningsprosjekt («Økologisk grønnsaksproduksjon») ble det utført forsøk med falskt såbed i gulrot og hodekål. Erfaringene med falskt såbed fra dette prosjektet gav positive resultater mht. tidsforbruk til manuell ugrasrenskning. I ett av forsøksåra ble det dog ikke store forskjeller mellom de ulike behandlinger. Dette året var det svært sen våronn. Konklusjonen fra prosjektet var at falskt såbed reduserer ugrasmengda og tida som trengs til manuell ugrasrensing, forutsatt at man har god nok tid om våren.
Strategien med falskt såbed er ganske fleksibel og man kan avbryte gjennomføringa dersom forholdene krever det. En annen fordel med falskt såbed er at tiltaket ikke medfører nevneverdige merutgifter. Ved bruk av falskt såbed er det også viktig å ta i betraktning at skade av ulike insekter kan variere med såtida, f.eks. gjør jordloppe mest skade når plantene er små.
Ved såing er det av stor betydning at man i utgangspunktet får så rask oppspiring av kulturveksten som mulig. Forsøk (figur 3.36) i Sverige har vist at om oppspiringen til korn (bygg) forsinkes, vil dette ha svært store konsekvenser for konkurranseforholdet mellom kulturveksten og ugraset, som i dette forsøket var dominert av meldestokk. Forsøket viste at tre dagers forsinket oppspiring doblet mengde ugras.
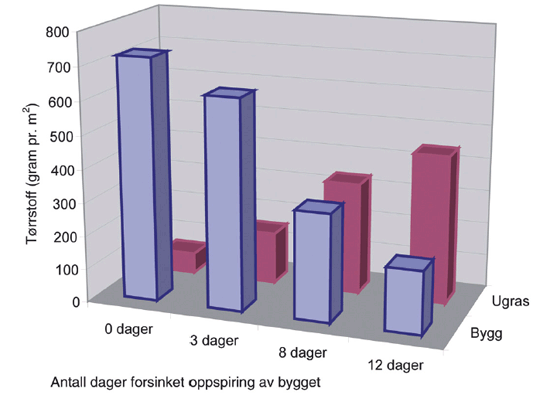
Flere ulike forhold i jorda, bl.a. jordtype og struktur, og jordmikroklimaet, herunder fuktighet og temperatur, påvirker oppspiringen til så vel kulturveksten som ugraset. I utgangspunktet kan vi likevel regne med at et godt tilberedt såbed vil gå i favør av kulturplanten. En annen viktig faktor er sådybden. For småfrøete arter sier det seg selv at man ikke må så for dypt. Ved dyp såing vil frøet bruke opp oppslagsnæringa og dø på vei opp. For storfrøete arter, som korn, står man friere til å bestemme sådybde. Dyp såing vil kunne medføre dårligere konkurranseevne mot ugraset, men det er viktig å tenke på at kornet skal ha fuktighet nok til å spire. Ved tilstrekkelig fuktighet vil grunn såing gi raskere oppspiring og bedre konkurranseevne. Riktig sådybde er så grunt som mulig uten at det skaper tørkeproblemer for frøet.
Ved innstilling av sådybde kan likevel andre faktorer også måtte tillegges vekt. I økologisk korndyrking er det for eksempel vanlig at man gjennomfører såkalt blindharving, dvs. at man harver rett før kornet viser seg på overflaten. Tanken med slik harving er at den skal drepe nylig spirt ugras, uten at kornet skades nevneverdig. Ved svært grunn såing vil kornet komme til overflaten før ugraset har spirt, og dette vil vanskeliggjøre blindharvinga. En dypere såing vil derfor øke selektiviteten mellom korn og ugras, slik at blindharvinga rammer ugraset, men sparer kornet. Ved dyrking av vekster som spirer sent, kan man utnytte det at ugraset spirer raskere enn kulturveksten. I for eksempel gulrot kan man utføre flamming (termisk kontroll) rett før gulrota spirer, slik at åkeren nærmest er fri for ugras den første tida etter at gulrota har spirt opp.
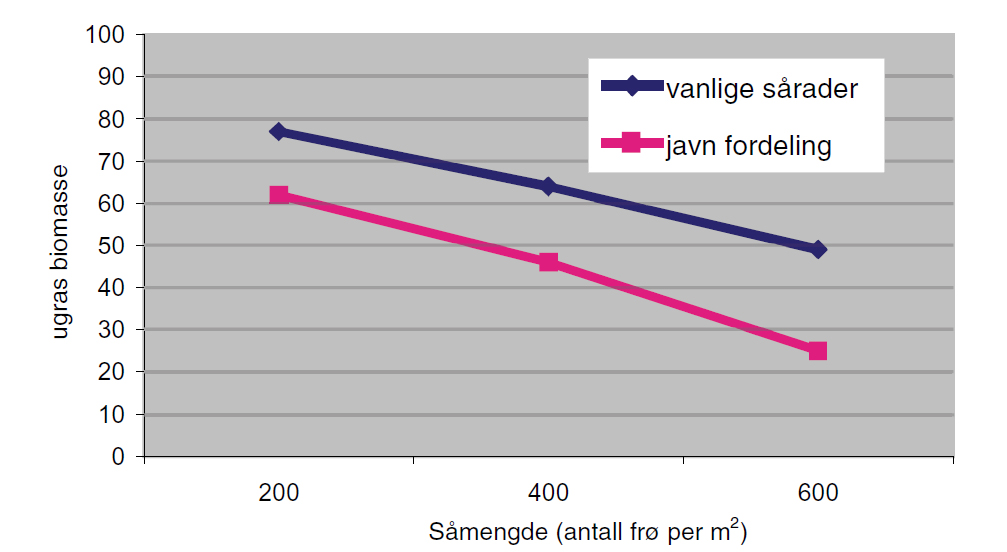
Ved økt plantetetthet vil kulturveksten konkurrere sterkere med ugraset, men intraspesifikk konkurranse øker også. Etter en tid vil kulturveksten konkurrere så sterkt med seg selv at avlinga går ned. Ved stigende såmengder vil prosentvis plantedød øke både for de minste kulturplantene og for ugraset, det siste avhengig av de ulike ugrasartenes konkurranseevne. I svenske undersøkelser er det funnet at en fordobling av såmengda i forhold til normalen kan medføre 30–60 % mindre ugrasbiomasse.
Hvis en kulturvekst er etablert, slik at frøet eller såkornet er optimalt fordelt utover, vil kulturvekstens konkurranseevne mot ugraset øke betydelig. Nylig publiserte resultater fra Danmark (Fig. 3.37) viser at man ved kombinasjonen økt såmengde til 600 frø per m2 og optimal frøfordeling, dvs. ikke ordinære sårader men «like langt mellom alle frø», reduserte ugrasbiomassen med 60 % og økte kornavlinga med 60 % sammenlignet med vanlig praksis, dvs. 400 frø sådd i rader. Både denne undersøkelsen og andre har vist at den optimale såmengda, både mht. virkning på ugras og avling, alltid vil være avhengig av hvor jevnt frøet fordeles på såflaten. Selv om disse resultatene virker ganske overbevisende, er ikke denne type såing lett å gjennomføre i praksis. Det kreves ny og kostbar såmaskin, og på visse jordtyper kan en slik utsåing også bety større problemer med skorpe (i en sårad spirer mange frø tett innpå hverandre og «løfter i flokk», mens frø som ligger spredt, må ta belastningen alene).
Sjukdommer og plantetetthet
Plantetetthet kan påvirke sjukdomsutviklingen bl.a. ved å påvirke mikroklimaet. Siden sjukdomsutvikling ofte fremmes av fuktige forhold, kan man tenke seg at et svært tett bestand vil øke «trivselen» for en rekke sjukdomsframkallende mikroorganismer. Likevel ønsker man å ha så dekkende bestand som mulig for å konkurrere ut en del ugras. Et tett ugrasbestand vil gjøre samme skade som tett kulturplantebestand med tanke på mikroklima, men vil kanskje hindre spredning av sjukdom fra plante til plante. Riktig sådybde og godt såbed vil også være av betydning for sjukdom. For dyp såing vil gi forsinket spiring, noe som også påvirker sjukdommer. Sammenhengen mellom plantetetthet og sjukdomsspredning er kompleks, og mange forsøk er utført uten at resultatene er entydige. Litt generelt kan man si at de mobile sjukdomsorganismene, eller de som spres lett i vekstsesongen, trives og spres ekstra godt i tette plantebestander. Mange har erfaringer med at for eksempel mjøldogg i korn gjør mindre skade og altså trives dårligere i litt tynnere åkre enn i tette, frodige åkre.
Skadedyr og plantetetthet
Betydningen av plantetetthet er lite undersøkt i forhold til skadedyr. Vi vet imidlertid at teger liker seg best i tynne bestander av f.eks. kålrot. Da kan det være en fordel å vente med tynning av direktesådde grønnsaker til etter at tegeangrep har avtatt. Å vente har også den fordelen at planter som er angrepet, kan tynnes bort.
Ugras og høstetid
Høstetida er nok ikke den aller viktigste faktoren når det gjelder konkurransen mellom kulturplante og ugras, men man kan se for seg en mer indirekte effekt: En tidlig høstet kultur kan gi bedre tid til mekanisk ugrasregulering om høsten. Et eksempel kan være stubbarbeiding med påfølgende pløying om høsten for å tyne kveka. Tidlig høsting kan også muliggjøre jordarbeiding mot ugraset før tilsåing av grønngjødslingsvekst eller fangvekst.
Sjukdommer og høstetid
Optimal høstetid avhenger av for eksempel valg av kultur, sort, såtid og ikke minst klima. Valg av vekster til modning gjøres ofte på grunnlag av ønske om høyest mulig avling. For mange kulturer vil de sene sortene gi større utbytte enn de tidlige. Lengre vekstsesong gir imidlertid større risiko for sjukdomsangrep. I tillegg risikerer man fuktigere vær utover høsten, noe som kan utsette høstingen enda lengre. For korn og potet ser man ofte økt angrep av en del sjukdommer ved sent høstetidspunkt.
Skadedyr og høstetid
Når et skadedyr angriper den plantedelen som skal være salgsvare, kan vi noen ganger unngå skade ved å høste tidlig. Et eksempel på dette er når vi ved hjelp av limfeller ser at det kommer et sterkt angrep av annen generasjon av gulrotflue. Registreringen er basert på voksne fluer før og under egglegging. Etter at larvene klekker ut av eggene, går det noen dager før larvene kommer seg inn i gulrøttene. Dersom røttene er store nok til å høstes, kan vi med fordel sette i gang med høsting av de ytterste radene (der angrepet er sterkest), og fjerne matgrunnlaget til skadedyret. Selv om gulrotavlinga blir noe mindre, vil den i hvert fall være salgbar og uten flueskade.
Arts- og sortsblandinger er ikke mye i bruk i dag, men ser vi litt i «glasskula», kan det her være mange elementer for utvikling av robuste dyrkingssystemer for framtida. Det er godt dokumentert at systemer med stor plantediversitet generelt er mer robuste i en plantevernsammenheng enn store flater med kun en kulturplante.
På mange måter vil arts- og sortsblandinger være i slektskap med det vi senere skriver om dekkekulturer. Bruk av dekkekultur vil si at vi sår inn en «ekstra» vekst som vi ikke skal høste, men som vi dyrker av andre grunner, mens vi for artsblandinger snakker om det å dyrke minst to forskjellige kulturvekster som skal høstes. Begge disse typene dyrkingssystem vil si at plantediversiteten øker, og mange av effektene vil kunne forklares ut fra de samme virkningsmekanismer. Et eksempel er insekter som blir forstyrret i vertsplantesøkingen når det dyrkes kombinasjoner av ulike vekster. Bruk av dekkekultur er mest aktuelt med tanke på skadedyr og ugras. Arts- og sortsblandinger er i tillegg svært interessant til kontroll av sjukdomsorganismer.
For å definere en artsblanding kan vi se for oss to ytterpunkter. På den ene siden har vi ensidig drift med store arealer dyrket med samme kulturvekst, for eksempel en åker med 100 dekar gulrot. I den andre delen av skalaen har vi systemer med større plantediversitet, for eksempel samplanting med gulrot og løk på annenhver rad. En mellomting mellom disse ytterpunkter kan være grønnsaker som dyrkes i 10 meter brede striper, med forskjellige grønnsaksarter side om side bortover åkeren.
Når det gjelder skadedyr, har mange undersøkelser vist at samplanting har en effekt. Det finnes lite dokumentasjon på effekten av «stripedyrking», men vi kan nok forvente oss en positiv effekt også her.
Artsblandinger med andre vekster enn grønnsaker kan være aktuelle, men mest realistisk er nok artsblandinger innen frukt, for eksempel eple og pære. Sortsblandinger er lite brukt i dag, men kan i framtida være aktuelt for flere kulturvekster. Korn og frukt er eksempler på kulturvekster hvor dette kan ha mye for seg. For begge gruppene er det hovedsakelig i relasjon til sjukdommer, for eksempel epleskurv, at sortsblandinger kan være interessant. Innen korn er dette en reise tilbake i tid. Før ble det ikke dyrket rene linjer, men en blanding av hva vi kan kalle ulike sorter. I dag er det nok mer aktuelt å komponere konkrete blandinger og blandingsforhold, og ikke bare bruke tilfeldige blandinger.
Selv om det er nærliggende å tenke sjukdomskontroll i relasjon til sortsblandinger, kan blandinger være interessant også i forhold til skadedyr og ugras.
Ugras
Flere grønnsakskulturer som løk, purre og gulrot har liten konkurranseevne mot ugraset. I grønnsakskulturer som såes sent, eller dekker jorda sent må det settes inn betydelige ressurser, det være seg mekaniske eller manuelle tiltak, for å kontrollere ugraset. I et sveitsisk prosjekt ble det undersøkt om man kunne kompensere for manglende konkurranseevne hos purre ved å dyrke den sammen med en annen og mer konkurransesterk vekst. Selleri ble valgt og dyrket i lag med purren i et «annenhver rad»-system (figur 3.38). Ulik tetthet av de to kulturene ble undersøkt, og samplanting av de to vekstene gav et system som var mer konkurransesterkt mot ugras enn purre alene. Ett av forsøkene viste at hvis man holdt jorda fri for ugras fram til 4 uker etter planting, og etter den tid lot ugraset vokse fritt, fikk man følgende avlingsreduksjoner sammenlignet med tilsvarende kultur(er) uten ugras: Purreavling redusert med 48 %, selleriavling redusert med 25 % og samplanting purre/selleri-avling redusert med 25 %. Et problem i disse forsøkene var at kvaliteten (størrelsen) på purren ble redusert ved samplanting. Et praktisk problem ved slike systemer er dessuten at samplanting kan vanskeliggjøre mekanisk ugrasrenhold.

Idéen om sortsblandinger i korn oppstod for å redusere angrepet av sjukdommer ved å bruke sorter med forskjellig resistensegenskaper. I tillegg er det funnet at sortsblandinger i korn har redusert ugrasforekomsten. Det er nærliggende å tro at dette skyldes to forhold: a) En frisk kornbestand, lite angrepet av sjukdommer, vil konkurrere bedre med ugraset enn hva tilfelle er for syke planter. b) Om sortene har litt ulike egenskaper mht. næringsopptak, morfologi og annet, vil den intraspesifikke konkurransen (mellom kornplantene) bli mindre, og blandingen vil kunne utnytte næring, vann og lys bedre enn en monokultur (bare en sort). En slik forbedret utnytting av vekstvilkårene vil antakelig også medføre forbedret konkurranseevne overfor ugraset.
Sjukdomsorganismer
Virkninger av arts- og sortsblandinger på forekomst av sjukdom på jordbruksvekster er særlig undersøkt i korn. I flere land i Europa arbeides det fortsatt aktivt med dette. Et sortsblandingssystem kan for eksempel omfatte 2–5 sorter med samsvarende modningstid og kvalitet, men med ellers ulike egenskaper. Allerede på 1800-tallet viste undersøkelser reduksjon i forekomst av rustsjukdommer i blandinger av hvete og havre. Også senere undersøkelser viser at blandinger av arter, sorter eller linjer (innen samme sort) av korn gir lavere angrep av sjukdommer som for eksempel rust, mjøldogg og sjukdommer i rot og stengelbasis. Lavere sjukdomsangrep i slike blandinger kan forklares ved hjelp av tre mekanismer: 1) Når man har sorts- eller artsblandinger, vil sjukdomsorganismen i første omgang oftere støte på blader den ikke greier å leve og oppformere seg på. Den første faktoren er derfor redusert spredning og oppformering av sjukdomsorganismen som følge av at mottakelig bladareal er mindre og ligger mer spredt. Spredningen videre fra de angrepne bladene blir i neste omgang også begrenset, både fordi det vil være lavere grad av oppformert smitte (sporer) enn i en monokultur, og fordi de sporene som er produsert, igjen vil ha mindre sjanse til å lande på en mottakelig plante pga. avstanden mellom disse. 2) De resistente plantene utgjør en fysiske barriere i en slik blandingsåker (figur 3.39). 3) Man kan få en «vaksineringseffekt» ved at sporer lander på en plante som ikke er vertsplante for sjukdomsorganismen. Soppsporene som lander, kan likevel sette i gang kjemiske eller fysiske forandringer i planten som gjør at den i visse tilfeller kan motstå et senere angrep av sopper som ellers ville ha framkalt sjukdom i den. Mekanismen bak dette kalles indusert resistens.
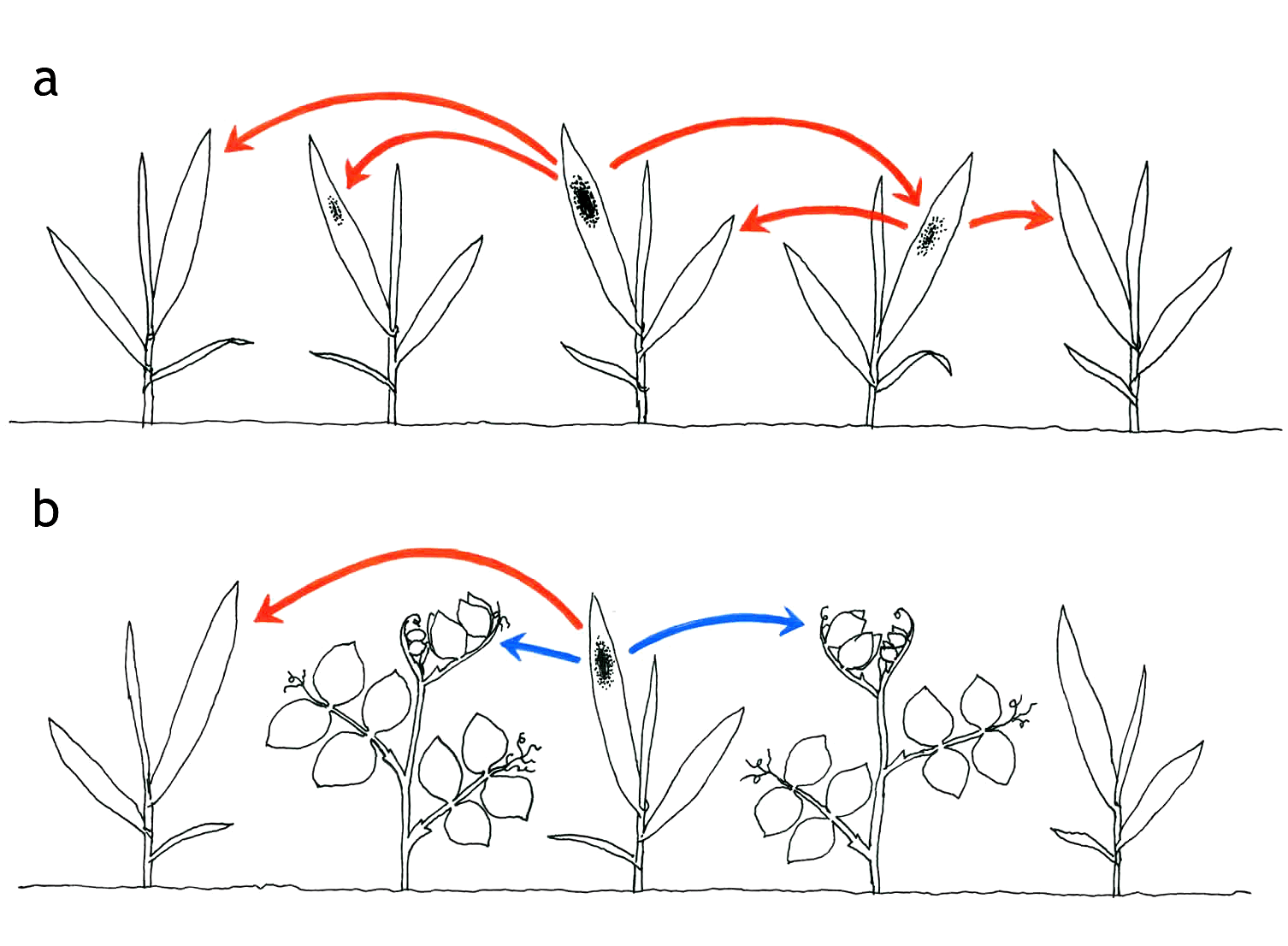
Samdyrking av erter og bygg er kanskje det eksemplet som er mest kjent i praksis her i landet. En dansk undersøkelse fra økologisk dyrking har vist at angrepet av mjøldogg i bygg var mye lavere der bygg var dyrket i blanding med erter, enn i bygg dyrket i renbestand.
Skal man for eksempel dyrke sorts- og artsblandinger av korn, er man avhengig av at sortene som inngår, modner på omtrent samme tid. Dette kan være en praktisk begrensning.
Skadedyr
Et klassisk eksempel på bruk av artsblandinger for kontroll av skadedyr er samplanting av grønnsaker. Blar man i eldre litteratur om økologisk grønnsaksproduksjon, trekkes ofte samplanting fram som et effektivt tiltak for å hanskes med skadedyrene. Flere vekster har i slik litteratur blitt lansert som gode kandidater for å holde skadedyrene på naboplanten borte, men det finnes få undersøkelser som underbygger påstandene. En vanlig kombinasjon som anbefales, er å dyrke løk sammen med gulrot for å holde gulrotflua på avstand. For denne kombinasjonen finnes det faktisk undersøkelser så langt tilbake som til 1930-årene, og flere av undersøkelsene har vist at gulrota blir mindre angrepet. Dessverre er ikke metoden funnet å være så effektiv (65 % mindre egglegging i en undersøkelse) at den har fått noe stort kommersielt omfang. Den brukes litt i småhager. I en del av litteraturen blir forstyrret vertsplantesøking (løken avgir aromatiske stoffer som forstyrrer gulrotfluas vertsplantesøking) trukket fram som hovedgrunnen til mindre angrep av skadedyr ved samplanting. For kontroll av skadedyr i frukt kan man tenke seg både å blande ulike fruktsorter, for eksempel eplesorter, og å blande ulike fruktarter.
Fangstplanter eller lokkeplanter er bevisst bruk av sorter eller arter som virker sterkt tiltrekkende på skadedyr, for å lokke dem vekk fra hovedkulturen. Fangsplanten kan være en annen art enn hovedkulturen, men den kan også være samme art, men en annen sort, eller til og med samme art og sort, men i et annet utviklingsstadium. Metoden er mest brukt i integrerte systemer der man sprøyter fangstplantene med et sterkt kjemisk skadedyrmiddel for å drepe skadedyrene, men den blir mer og mer brukt også i økologisk dyrking, der man ødelegger fangstplantene og dreper skadedyrene på den måten.
Bruk av fangstplanter er best kjent fra radkulturer som potet og grønnsaker, men det finnes også klassiske eksempler fra bomull og lusern. I Norden er bruk av fangstplanter mest undersøkt som en metode til å redusere angrep av gulrotsuger. Da brukes gulrot som er sådd tidligere enn hovedkulturen, enten i potter eller direkte i jorda. Fangstplantene står som en ramme rundt hovedkulturen og fanger opp voksne sugere under innflyvningen til åkeren (jfr. figur 3.40). Når hovedinnflyvningen er over, freses fangstplantene ned.
Et opplegg der mainepe blir brukt som fangstplante, er undersøkt i blomkål og kålrot i Norge. Mainepa tiltrekker både kålflue og nepejordlopper. Forsøkene viste at når mainepe var plantet inn i hovedkulturen (jfr. figur 3.40) ble nepeplantene fullstendig ødelagt av jordlopper, mens nyutplanta blomkål var nærmest urørt. Tilsvarende resultat med mainepe som fangstplante fikk man i kålrot: Nepene ble oppspist av kålflua, mens kålrota fikk være i fred.
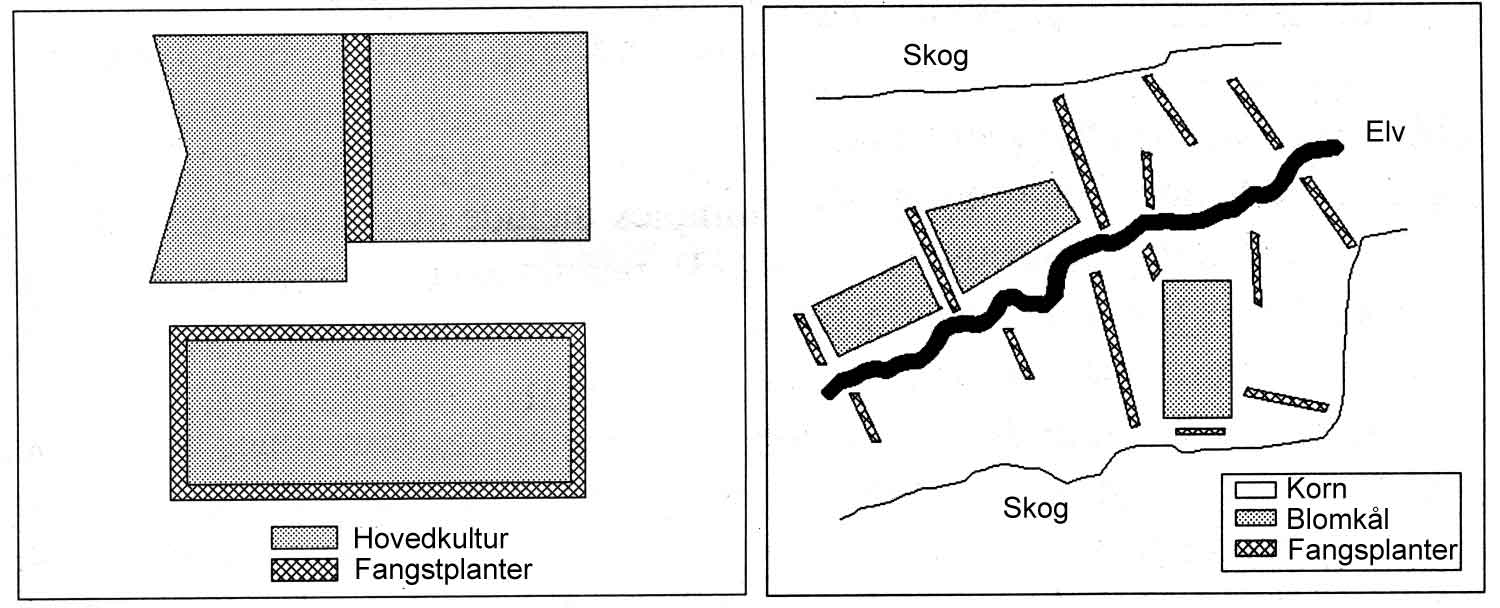
Fangstplanter er foreslått brukt i kombinasjon med andre metoder som f.eks. underkultur, samplanting, insektgjerder og repellenter for å «dytte» insekter vekk fra hovedkulturen og samtidig dra dem til fangstplanter.
Oppsummering, arts- og sortsblandinger
Ved økologisk dyrking vil det ofte være behov for å sette inn flere tiltak i en mer sammensatt strategi mot en bestemt skadegjører. Fordi bruk av arts- og sortsblandinger ofte ikke vil være effektive nok til å løse plantevernproblemet alene, vil det være mest realistisk å kombinere denne metoden med andre tiltak.
Underkultur i korn (figur 3.44) er nok det vanligste eksemplet på praktisk utnyttelse av dekkekultur i Norge. Dekkekultur i frukt (figur 3.44) er ikke så vanlig, og når det gjelder grønnsaker (figur 3.44), er bruken foreløpig mest knyttet til forskning. Underkultur i korn har sitt utspring i ønsket om å ha en nitrogenfikserende vekst i omløpet. Hvis en ikke-belgvekst som for eksempel raigras, benyttes som underkultur i korn, vil ofte hensikten være fangvekstrelatert, d.v.s. redusere faren for erosjon og utvasking. Selv om hovedhensikten med en underkultur ofte er relatert til næringsforsyning, vil underkulturen i større eller mindre grad også påvirke ulike skadegjørere. I korn vil nok effekten på ugraset være det mest iøynefallende. I andre kulturvekster, som frukt og grønnsaker, er det også godt dokumentert at dekkekulturen kan ha gunstig effekt på skadedyrsituasjonen. Selv om det tradisjonelt har vært mest fokus på effekter på ugras og skadedyr, vil en dekkekultur også kunne ha betydning for ulike sjukdomsorganismer.
Bruk av dekkekultur vil, i tillegg til det som allerede er nevnt, også påvirke mange andre faktorer. Selv om dekkekulturen kan ha mange fordeler, er bruken (bortsett fra i korn) likevel ganske begrenset. Det er flere grunner til den begrensede bruken, men konkurranse med kulturplantene er sannsynligvis den viktigste årsaken. Egenskapene til de ulike grupper av kulturvekster, ikke minst konkurranseevna og måten de dyrkes på, gjør at det er vanskelig å gi generelle oppskrifter på bruk av dekkekultur i praksis. Inntil videre er nok bruk av dekkekultur mest aktuelt i korn, kanskje også i frukt- og bærvekster.
Ugras og dekkekultur
Hvis en dekkekultur etableres på en god måte, blant annet at flerårige ugras bekjempes før etablering av dekkekulturen, kan den være et effektivt tiltak mot ugraset. En levende dekkekultur vil generelt ha mye bedre effekt på ugraset enn et dødt plantedekke. Dette ble tydelig vist i en amerikansk undersøkelse hvor det viste seg at man måtte ha tredobbel mengde dødt plantemateriale (tørrstoff) av lodnevikke for å få samme virkning på ugraset som et levende bestand av denne belgveksten (tekstboks 3.12).
Et levende dekke påvirker lysforholdene mye mer enn et dødt dekke ved at:
Levende og dødt plantemateriale vil gi ganske lik effekt på temperaturen i jorda. Maksimumstemeraturen synker og minimumstemperaturen stiger, d.v.s. mindre temperaturvariasjon gjennom døgnet. Fordi frøene til flere ugras trenger en viss temperaturvariasjon for å spire, fører dette til mindre spiring.
Andre effekter:
Hvilke av faktorene som påvirker ugraset mest, vil variere for ulike typer plantedekke, men lys, dernest temperatur, er sannsynligvis viktigst.
Skadedyr og dekkekultur
En stor del av forskningsinnsatsen innen bruk av dekkekultur (underkultur) i radkulturer har dreid seg om å redusere angrep og skade av insekter. Mange av studiene har dreid seg om kombinasjonen korsblomstrete vekster (kålvekster) med en kløverart brukt som underkultur. I flere tilfeller er det funnet at dekkekultur har redusert skadedyrproblemene. Ulike virkningsmekanismer er foreslått for å forklare effektene som er funnet (tekstboks 3.13).
En av de som har arbeidet mest med å avdekke virkningsmekanismer er den britiske skadedyrforskeren S. Finch. Han og hans medarbeider stiller seg kritiske til en del av hypotesene som er nevnt i figur 3.41 og har kommet med en egen teori som de har kalt « passende/ikke-passende landinger» (figur 3.42) av skadeinsekter på korsblomstrede planter. Denne teorien bygger på langvarige og detaljerte studier av hvordan ulike insekter oppførerer seg i ulike situasjoner. Punktvis kan teorien settes opp som følger (figur 3.43):

Dekking av jorda med et dødt dekkemateriale vil påvirke de ulike typer skadegjørere i større eller mindre grad. Metoden er mest aktuell i radkulturer som grønnsaker, frukt og bær. Aktuelle materialer som kan brukes til slik jorddekking, inkluderer både industrielle produkter som plast og papir, og organiske materiale som halm, gras- eller kløverkapp, sagflis og gjødsel eller kompost. Dødt materiale kan i stor grad også påvirke kulturplantene gjennom gjødselvirkning, effekter på lokalklima og lignende. Den totale virkningen blir dermed summen av effekter, inkludert virkningen på skadegjørere og andreforhold som påvirker planteveksten.
Under varme og tørre klimaforhold (tropiske og subtropiske) vil dødt materiale ofte medføre enda flere fortrinn, for eksempel økt jordfuktighet og tilførsel av organisk materiale til jorda. I tempererte klimasoner derimot, kan effekten av lavere jordtemperatur, hvilket en del dekkmaterialer fører til, være uheldig.

I tekstboks 3.14 har vi oppsummert ulike effekter av dødt materiale på ugras, plantesjukdommer, insekter og edderkoppdyr. I tekstboks 3.15 vil du finne noen eksempler på bruk av metoden i grønnsaker og frukt.
Det kan endre jordas struktur, fuktighet og temperatur, lysforhold ved jordoverflaten og plantenes tilgang på næringsstoffer. Alle disse faktorene kan ha innvirkning på planteveksten som videre påvirker plantenes konkurranseevne mot ugras, toleranse mot skadeinsekter og motstandsdyktighet mot sjukdommer.
Forskjellige typer materiale kan hindre ugrasfrø i å spire og frøplanter i å etablere seg ved å:
I et naturlig miljø vil det være et samspill mellom mikroklima, fysiske barrierer og kjemiske effekter, og det er derfor vanskelig å skille ut betydningen den enkelte faktor har på frøspiring og vekst av frøplanter.
Når det gjelder insekter og edderkoppdyr, vil jorddekking medføre en forandring av agroøkosystemet som vil påvirke både kulturen og skadegjøreren så vel som naturlige fiender. At jorddekkingssystemer har en slik kompleksitet, gjør allmenngyldige konklusjoner vanskelig, men noen generelle linjer kan likevel trekkes:
Jorddekking kan forstyrre insektenes vertsplantevalg ved at plantetettheten forandres, naturlig forekommende kjemikalier tiltrekker eller frastøter insekter eller den endrede jordbunnsfargen (i forhold til svart jord) kan ha en effekt.
Innflytelsen dødt materiale har på ugras og insekter, vektlegges ofte mest, men slik jorddekking kan også i stor grad påvirke forekomst og utvikling av sjukdommer ved å:
Grønnsaker
Særlig i produksjonen av tomater, har flere forsøk med dekkmateriale vist at svart plast hemmer ugraset effektivt. Papir er et annet dekkmateriale som kan brukes i grønnsaksproduksjonen, men et problem med papir er at det blir raskt brutt ned, og effekten mot ugraset blir dermed dårligere. Papir er likevel velegnet i vekster som har kort vekstsesong, slik som salat. En fordel med papir er at man ikke trenger å fjerne det etter høsting, fordi det etter hvert går i oppløsning og forsvinner av seg selv. Det finnes papir av forskjellig kvalitet på markedet, for eksempel papir som er behandla med voksfilm, basert på oljeekstrakt, for å forsinke nedbrytingen. Forsøk har vist at behandla papir kan hemme ugrasveksten i mer enn 10 uker, mens ubehandla papir holder i 6–9 uker.
En annen mulighet er å dekke jorda med plantemateriale, det kan være halm eller opphakkede belgvekster fra et naboskifte på gården. En ulempe med det siste er at en slik praksis beslaglegger store arealer for å få nok opphakka plantemateriale. Mengda organisk dekkmateriale som kreves for en effektiv ugrasbekjempelse, avhenger av flere faktorer, for eksempel sammensetningen av ugrasfloraen, hvor smått ugraset er ved pålegging, hvor raskt plantematerialet brytes ned og hvor sterk konkurranseevne kulturveksten har mot ugraset. Det må for eksempel legges på mer avklipp i gulrot enn i hodekål. Siden det er så mange faktorer som påvirker hvor mye organisk dekkmateriale som kreves, er det vanskelig å komme med generelle tilrådinger om hvor mye avklipp som behøves. Det vi likevel kan si sikkert, er at man minst må ha en mengde på 600 g tørt materiale per m2, dette tilsvarer ca. 4,5 tonn friskmasse per dekar. En måte å redusere behovet per dekar på, er å dekke jorda kun mellom plantene i rada, og bekjempe ugraset mekanisk mellom radene. Mange typer dødt materiale vil ikke ha tilfredsstillende virkning på flerårige ugras. Slikt ugras bør derfor fjernes før dekkmaterialet blir lagt på.
Som allerede nevnt, har ulike dekkmaterialer også effekt på skadeinsekter. I forsøk i gulrot har dekking med sagflis og avklipp av belgvekster blitt testet mht. effekter på gulrotsuger. Forsøkene viste at disse dekkmaterialene kan redusere skaden betydelig. De samme undersøkelsene viste også at dekking gav større tetthet av visse nyttedyrarter. Utenlandske forsøk har vist at reproduksjonen av planteparasittære nematoder, som for eksempel rotgallnematoder på tomat, varierer ved bruk av plastdekke med forskjellig farge. Bruk av reflekterende dekkmateriale har også forsinket utviklingen av mosaikkvirus i squash som en følge av reduksjon i bladluspopulasjonen. Bladlus fungerer som vektor i overføringen av mange virussjukdommer. Flere undersøkelser har vist at insektspopulasjoner er blitt redusert betraktelig ved bruk av plast (eller annet materiale) med visse farger.
Fruktproduksjon
Jorddekking kan være fordelaktig i fruktproduksjonen, særlig i økologisk landbruk. Svart plast hemmer ugrasutviklingen effektivt i frukt. For å øke plastens holdbarhet er det fordelaktig å benytte vevd plast. Bark, trekutt eller et kompakt lag av halm kan også være velegnede dekkmaterialer i frukt, men et lag på minst 10–15 cm er nødvendig for å gi langvarig effekt mot ugraset. Nedbrytbart materiale som bark, hvilket ofte er næringsrikt, har vist seg å være utilstrekkelig i ugraskontrollen.
Jorddekking i frukthagen har også hatt effekt på andre skadegjørere. svart plast reduserte for eksempel populasjoner av Pythium ultimum i den øvre rotsonen.
Til tross for at jorddekking har positive effekter i frukt, finnes det også ulemper, bl.a. kan det være et problem å kontrollere ugraset som vokser i kantene av dekkematerialet. Et annet problem er ugras som vokser inntil stammen av frukttrærne. Det må som regel håndlukes. Jordrotter kan også være et problem ved jorddekking i frukthagen fordi tett dekke skaper et ideelt miljø for rottene. Type dekkmateriale ser ut til å ha betydning for jordrotteproblemet, og ødeleggelse av frukttrær har i enkelte studier vist seg å forekomme sjeldnere ved bruk av trekutt enn ved bruk av andre dekkmaterialer. Som nevnt tidligere, kan problemer med jordbårne sjukdommer reduseres ved bruk av dødt materiale, men motsatte effekter er også kjent, for eksempel økte rotsjukdommen Phytophthora ved bruk av halmdekke.
Aamisepp, A. og B. Wallgren 1979: Ogräs i stråsäd. Verkan av kemisk ogräsbekämping och andre odlingsåtgärder, 1950–1978. Aktuelt från lantbruksuniversitetet 437/438. Mark • Växter, Uppsala, 70 s.
Alkämper, J. 1976: Einfluß der Verunkrautung auf die Wirkung der Düngung. Pflanzenschutz-Nachrichten Bayer, 29, s. 191–235
Baumann, D.T., L. Bastiaans og M.J. Kropff 2001: Competition an crop performance in a leek-celery intercropping system. Crop Science 41 (3), s. 764–774
Baumann, D.T., L. Bastiaans og M.J. Kropff 2001: Effects of intercropping on growth and reproductive capacity of late-emerging Senecio vulgaris L., with speciel reference to competition for light. Effects of intercropping on growth and reproductive capacity of late-emerging Senecio vulgaris L., with speciel reference to competition for light. Annals of botany 87 (2), s. 209–217
Bostrøm,U. og H. Fogelfors 1999: Type and time of autumn tillage with and without herbicides at reduced rates in southern Sweden: 2. Weed flora and diversity. Soil and Tillage Research 50 (3–4), s. 283–293
Brandsæter, L.O. 1998: «Nattarbeiding», underkultur og falsk såbed. Fagseminar 11. og 12. februar 1998. Grønn Forskning 6, s. 8–12
Brandsæter, L.O. og M. Helgheim 2002: Nye belgvekstarter – potensiale for ugraskontroll og grønngjødsling i grønnsaker (Plantemøtet 2002). Grønn Forskning 2, s. 153–157
Brandsæter, L.O., J. Saur, A.K. Bakken, T. Wikmark og T. Fjeld 2005: Jordarbeidings-metoder for korndominerte dyrkingssystem – effekt på flerårige ugras. Grønn kunnskap 9(2): 368–374.
Coombes, D.S. og N.W. Sotherton 1986: The dispersal and distribution of polyphagous predatory Coleoptera in cereals. Annals of Applied Biology 108, s. 461–474
Dock Gustavsson, A-M. 1992: Feltforsøk med åkertistel. 33. svenska vaxtskyddsnotiser, ugras og ugrasbekampning, s. 73–77
Esbjerg, P., J. Jørgensen, J.K. Nielsen, H. Phillipsen O. Zethner og L. Øgaard 1983: Integreret bekæmpelse af skadedyr: med gulerødder; gulerodsfluen (Psila rosae F., Dipt. Psilidae) og ageruglen (Agrotis segetum Schiff., Lep., Noctuidae) som afgrøde-skadedyr model. Tidsskrift for Planteavl 87, s. 303–355
Finch, S. og M. Kienegger 1999: Host-plant finding by insects – «appropriate/ inappropriate landings» a mechanism based on the behaviour of pest insects of cruciferous plants. IOBC/WPRS Bulletin 22, s.157–161
Finch, S. og R.H. Collier 2000: Host-plant selection by insects – a theory based on ‘appropriate/inappropriate landings’ by pest insects of cruciferous plants.
Entomologia Experimentalis et Applicata 96, s. 91–102 Fykse, H. 1974: Studium av åkerdylle. II Utbreiing i Noreg, vokster og kvile – dels jamført med nærståande arter. Forskning og forsøk i landbruket Bind 25, s. 389–412
Fykse, H. 1993: Ugrasituasjonen i vårkorn ved ulike bekjempningsstrategiar. Informasjonsmøte i plantevern 1993, Faginfo Nr. 3 s. 172-180
Fykse, H. og H. Sjursen 1992: Forelesninger i herbologi: II. Rådgjerder mot ugras. Ås–NLH, 107 s.
Fykse,H. og K. Wærnhus 1997: Innverknad av såtida på utviklinga av ugraset og behovet for sprøyting i haustkorn. Grønn Forskning 2, s. 117–124
Griepentrog. H.W., J. Weiner og L. Kristensen 2000:. Increasing the suppression of weeds by varying sowing parameters. I : Proceedings from 13th International IFOAM Scientific Conference (Red. Alføldi. T., W. Lockeretz og U. Niggli), 173 s.
Hokkanen, H.M.T. 1991:Trap cropping in pest management. Annual Review of Entomology 36, s. 119–138
Holmegaard, J. 1987: Grøngødning og eftergrøder. Skarv, Holte, Danmark, s. 224.
Håkansson, S. 1979: Grunnläggande växtodlingsfrågor. II. Faktorer av betydelse för planteetablering, konkurrens och produktion i åkerns växtbestånd. Sveriges lantbruksuniversitet, Institutionen för växtodling, Rapport 72, Uppsala, ??s.
Håkansson, S. 1988: Competition in stands of short-lived plants. Density effects measured in three-component stands. Sveriges lantbruksuniversitet, Crop Production Science 3, Uppsala, 181 s.
Håkansson, S. 1995: Ogräs och odling på åker. Aktuelt från lantbruksuniversitetet 437/438. Mark • Växter, Uppsala, 70 s.
Håkansson S. 2002: Soil tillage and weeds on arable land. I: Weed Science Compendium 2002 Edition (Red. Jensen, J.E., J.C. Streibig og C. Andreasen). Samfundslitteratur KVL-bogladen, København (DK), 476 s. Levenfors, J., J. Lager og B. Gerhardson 2001: FAKTA Jordbruk Nr.1 2001 (utgis av SLU)
Rasmussen, K. 2000: Can slurry injection improve selectivity of weed harrowing in cereals? 4th Workshop of the EWRS working group Physical Weed Control, 20–22 March 2000, Elspeet, Nederland.
Rouhiainen, S.K., J. Vaaisanen, P. Vanhala og T. Løtjønen 2003: Mid-summer bare fallow effective in controlling perennial weeds. Nordic Association of Agricultural scentists 22nd Congress, July 1–4 2003, Turku, Finland.
Suhr, K., J. Thejsen og K. Thorup-Kristensen 2005. Grøngødning, efterafgrøder og dækafgrøder. Dansk Landbrugsrådgivning. Landcenteret, Landbruksforlaget, Århus, 264 s.
Tobiasson, M. og G. Danielsberg 2004: Teknikk og strategier mot ugras: Brakking og pussing. Grønn kunnskap (Jord- og plantekultur 2004) 8 (1), s. 368–380
Ögren, E. 1998: Kväve från gröngödsling måste kunna styras. Uppsala, Sveriges lantbruksuniv., Fakta. Trädgård, 1998: 8, 2s.
Ögren, E. , B. Båth og B. Rämert 1998: First and second year nitrogen effects of autumn and spring-incorporated green-manure crops in field vegetable production. Swedish Journal of Agricultural Research 1998, v. 28(3), s. 137–146
Uvah, I.I.I. og T.H. Coaker 1984: Effect of cropping on some insect pests of carrots and onions. Entomologia Experimentalis et Applicata 36, s. 159–167
Viken, T.H. 2003: Ikke-vertsplanters innvirkning på klekking av potetcystenemtoden Globodera rostochiensis. Hovedoppgave ved Norges landbrukshøgskole / Planteforsk Plantevernet, 64 s.
Wivstad, M. 1989: Försök vid försöksavdelningen för växtföljder. Ettårig gröngödsling. Alternativodlingsbrevet Maj 1989, (nr 16), s. 4–8
Selv om forbyggende tiltak skal være fundamentet for kontroll av skadegjørere ved økologisk dyrking, vet vi at direkte tiltak ofte vil være helt nødvendig for å lykkes med produksjonen. Ikke minst gjelder dette for kontroll av ugras i mange kulturvekster. I dette kapittelet vil vi komme nærmere inn på biologisk kontroll, fysisk og termisk kontroll, og bruken av såkalte alternative plantevernmidler. Mye av det vi omtaler i dette kapitlet, for eksempel ulike typer utstyr for ugraskontroll, er det som er tilgjengelig i dag, men litt informasjon om metoder som er under utvikling, er også tatt med.
Direkte tiltak kan vi definere som tiltak som rettes mot en skadegjører i den hensikt å drepe skadegjøreren eller redusere tilvekst og formering. Dette høres i utgangspunktet ganske greit ut, men i praksis kan det være vanskelig å sette klare skille mellom forebyggende og direkte tiltak. Intensjonene med de to tiltaksformene er imidlertid forskjellige: Mens forebyggende tiltak har som mål å hindre at en organisme skal utvikle seg til å bli et problem – en skadegjører, er målet med direkte tiltak å redusere omfanget av en organisme som alt er blitt en skadegjører, så mye at den er til å leve med.
Her har vi valgt å klassifisere de direkte tiltakene på følgende måte:
Biologisk kontroll er aktuelt både mot ugras, sjukdomsorganismer og skadedyr. Fysisk og termisk kontroll er på den andre siden mest forbundet med ugras. Imidlertid vil fysisk utestengning av skadedyr ved bruk av gjerde, fiberduk/ insektsnett eller tildekking av jordoverflaten også kunne kalles en fysisk metode. Selv om enkelte kanskje vil si at termiske tiltak (oppvarming) mot sjukdommer og skadedyr for sanering av skadegjørere i frø, småplanter, settepoteter m.m. hører hjemme under forebyggende tiltak, er dette tatt med her som et direkte tiltak.
Plantevernmidler er slett ikke det man i første omgang forbinder med kontroll av skadegjørere i økologisk landbruk. Regelverket tillater likevel noen stoffer, for eksempel svovel, for kontroll av epleskurv. Dessuten vil det helt sikkert i tida framover bli reist spørsmål om behov og ønske om nye «grønne» plantevernmidler. Mot visse vanskelige skadegjørere mener mange at slike midler i alle fall foreløpig er eneste farbare vei for å oppnå sikker produksjon. Bruk av naturlig forekommende signalstoffer, for eksempel feromoner, vil bare være aktuelt mot skadegjørere som aktivt søker etter vertsplanter, dvs. mot skadedyr. Vi har valgt å omtale denne type planteverntiltak som bruk av alternative plantevernmidler. Siden grenseoppgangen mellom ulike typer av plantevernmidler er ganske vanskelig, har vi noen ord om dette i en egen tekstboks (se tekstboks 4.1).
Betegnelsen plantevernmiddel og relaterte begreper brukes i ulike sammenhenger også i forbindelse med økologisk landbruk, blant annet i lover og regelverk og i utrednings- og forskningsdokumenter, så vel nasjonalt som internasjonalt. Ikke alltid blir begrepene like klart definert. Betegnelsene «syntetiske» og «kjemisk/syntetiske» plantevernmidler brukes ofte i nasjonale og internasjonale regelverk, men er heller ikke der nærmere definert. Man kan si at denne mangel på klare definisjoner delvis kompenseres ved at regelverket inneholder spesifikk informasjon om hvilke midler som er godkjente. IFOAM har i den forbindelse også definert kriterier for godkjenning av ulike produkter som kan brukes i plantevernsammenheng.
Plantevernmiddel av naturlig opphav er et nyere begrep som antyder at det virksomme stoffet har en naturlig opprinnelse, og dermed også er styrt av naturlige reguleringsmekanismer. Begrepet alternative plantevernmiddel omfatter vanligvis de stoffene som ikke er framstilt syntetisk. Også begrepet biopesticider defineres på ulike måter, enten at dette begrepet bergrenses til mikroorganismer som benyttes som plantevernmiddel, eller jfr. OECD-definisjonen hvor biopesticider også inkluderer feromoner, insekt- og plantevekstregulatorer, planteekstrakter, transgene planter og makroorganismer.
Biologisk kontroll defineres som bruk av levende organismer til å bekjempe planteskadegjørere. Biologisk bekjempelse av skadedyr så vel som sjukdomsorganismer og ugras deles vanligvis inn i tre prinsipielle metoder:
Introduksjonsmetoden
omtales ofte som klassisk biologisk bekjempelse, og omfatter introduksjon og etablering av nye arter i en region der de opptrer som fiender av skadegjøreren. Metoden brukes mot en ny skadegjører som har etablert seg, og som stammer fra et helt annet område. Det opprinnelige utbredelsesområdet ligger ofte langt borte, gjerne på et annet kontinent. I slike tilfeller er de lokale, naturlige fiender ikke tilpasset til å leve på eller av den nye skadegjøreren. Da kan en innføre naturlige fiender fra området hvor skadegjøreren opprinnelig hører hjemme. Ofte innføres få individer av en bestemt art. Blir etableringen vellykket, vil nytteorganismen relativt raskt formere seg og redusere bestanden av den nye skadegjøreren til akseptabelt nivå. Introduksjonsmetoden må brukes med forsiktighet fordi introduksjon av nye arter kan få uønskete økologiske konsekvenser, og true biodiversiteten i det nye området.
Oversvømmelsesmetoden
kan sammenlignes med bruk av et plantevernmiddel som gir en rask og direkte virkning på skadegjøreren (brukt som et biopesticid). Nytteorganismene masseoppformeres og slippes ut i store mengder. Metoden har kortvarig virkning, og nye individer må introduseres etter en viss periode. Alle arter av nytteorganismer som selges til bruk i biologisk bekjempelse, må her i landet være godkjent av Mattilsynet. De må tilfredsstille kravet til ønska agronomisk virkning, og ikke innebære noen risiko for uønska økologiske konsekvenser .
Ved konservering
skjer det ingen kunstig oppformering av nytteorganismene, men forholdene legges til rette for at de naturlige fiendene som finnes på stedet, får best mulige leveforhold slik at de kan bekjempe skadegjørerne.
Ved biologisk kontroll av ugras blir en levende organisme sprøytet eller satt ut i den hensikt å redusere ugras til et nivå som ikke er skadelig for kulturplantene. Effekten av nytteorganismen kan være direkte ved at viktige plantevev og funksjoner ødelegges slik at ugraset drepes eller hemmes i veksten. Nytteorganismen kan også ha en indirekte effekt ved at ugraset stresses nok til at det taper konkurransen med kulturplantene om ressursene. Levende nytteorganismer som har blitt undersøkt, eller er benyttet for biologisk bekjemping av ugras, kan deles inn i nyttedyr (insekter, midd og nematoder) og nyttemikroorganismer (bakterier, sopp og virus).
Prinsippet for selektiv ugraskontroll baserer seg på at nytteorganismene har utviklet seg og tilpasset seg et bestemt eller nært beslektede ugras gjennom tusenvis av år. Vi sier at slike organismer er vertsspesifikke. Nytteorganismene er ofte så sterkt tilpasset og spesialiserte at uten tilgang på vertsugraset sitt, vil de dø. Noen lever utelukkende av å beite på planten, mens andre parasitterer eller forårsaker vekstforstyrrelser og sjukdommer.
Noen av de først beskrevne suksessforsøkene innen biologisk ugrasbekjempelse fant sted i det nordøstre Australia hvor den innførte kaktusslekten Opuntia var blitt en stor plage mange steder ved at den ødela store beiteområder. På 1920-tallet ble den søramerikanske kaktusmøllen, Cactoblastus cactorum, sluppet løs. I løpet av noen år ble kaktusplagen kraftig redusert, takket være spredningen av møllen til nye områder. Man har anslått at kaktusene har blitt fortrengt fra jordbruksområder tilsvarende 25 millioner hektar. Dette er et eksempel på introduksjonsmetoden, klassisk biologisk kontroll.
Etter oppdagelsen av de første kjemiske sprøytemidlene, falt interessen for biologisk kontroll betraktelig. Først da fokus ble satt på alle ulempene ved bruken av kjemikalier, fattet forskere igjen interesse for biologisk kontroll. De første forsøkene skjedde etter prinsippet om at den sterkeste av flest mulig testete kandidater ville overleve. Denne ukritiske utsettelsen førte til nye plager i land som USA, Australia og New Zealand, ettersom «nytteorganismene» ofte skiftet vert eller angrep sårbare planteslag. Nå er det derfor strenge restriksjoner på utsettelse av nytteorganismer for bruk i biologisk kontroll av ugras, og et stort register av mulige vertsplanter må testes grundig på forhånd. De mest interessante kandidatene for biologisk kontroll samles inn og undersøkes for diett og overlevelsesevne. I tillegg må tidkrevende spesifisitetstester utføres for at ikke nært beslektede planter, som viktige kulturplanter og planter som har fysiske og kjemiske likheter, utsettes for mulige angrep fra nytteorganismene.
Nyttedyr til bekjempelse av ugras (insekter og nematoder)
I Norge er det ikke arbeidet spesielt med nyttedyr til bekjempelse av ugras. I USA derimot selges flere typer nytteinsekter gjennom firmaer som har spesialisert seg på oppformering av insekter. Det er for eksempel flere nyttedyr på markedet mot ugras som åkertistel, nikketistel, ulike knoppurter, stjerneknoppurt, prikkperikum, landøyda, lintorskemunn, giftkjeks (skarntyde), veivortemjølk og kattehale. For å bedre effekten brukes ofte flere typer nyttedyr, som har spesialisert seg på ulike organer av planten, sammen, slik at de rammer ugraset flere plasser og ikke konkurrer med hverandre. Nematoder er velkjente skadegjørere på planter, og noen er meget artsspesifikke. De har blitt lite undersøkt med tanke på biologisk kontroll av ugras, men det er ikke utenkelig at disse organismene utgjør et stort framtidig potensiale.

Nyttemikroorganismer til bekjempelse av ugras
Ugras er vertsplanter for en rekke virus som kan tenkes brukt i ulike strategier for ugrasbekjempelse. Virus har den fordelen at de kan frysetørres og være aktive selv etter flere års lagring. De kan dyrkes opp og ekstraheres fra planter som tolererer virusinfeksjonen. I USA pågår det forskning på interessante virus for bruk i biologisk ugrasbekjempelse.
Sjukdomsframkallende bakterier kan også brukes til biologisk kontroll av ugras. Bakterier har den fordelen at de er lette å formere, tåler frysing og i mange tilfeller også frysetørking. Det største hinderet for bruken av bakterier er at de ikke kan trenge aktivt inn i plantevevet på egen hånd. Dette kan løses ved å tilsette vætemidler som bryter vannspenningene i hulrommene i bladene, eller såring av planteoverflaten, slik at bakteriene finner inngangsporten. Mest kjent er kanskje bruken av bakterier på golfbaner for å kontrollere tunrapp. Bakteriene påføres i forbindelse med at graset klippes, slik at bakteriene trenger inn i kutta blad og stengler og tetter transportvevet.
Sopp er den gruppen av mikroorganismer som har blitt mest undersøkt i forbindelse med biologisk ugraskontroll. Sopp har vært brukt i klassiske kontrollstrategier. I den senere tid har det imidlertid blitt arbeidet mest med å isolere naturlig forekommende organismer på den aktuelle ugrasart. Disse formeres på kunstig medium og sprøytes ut (etter oversvømmelses-metoden som et bioherbicid) på samme måte som vanlige ugrasmidler. Fordelen er at soppen allerede finnes i miljøet, og forekomsten blir raskt redusert når tilgangen på vertsplanter blir mindre. Sopp er lett å dyrke i store mengder på kunstig medium, men ulempen er at de ofte krever optimale forhold for å infisere vertsugraset, det vil si 20-25 oC og 100 % luftfuktighet. Sprøyting i solskinn kan drepe sporene direkte, og indirekte ved at sprøytevæska på planteoverflata tørker for raskt opp. De lave nattetemperaturene som ikke er uvanlig i Norge om våren, vil også hemme soppsporene i å spire, selv om sprøyting på kveldstid kan gi gunstig luftfuktighet. Biologisk bekjempelse av åkertistel med bruk av både sopper og insekter har vært mye studert rundt i verden. I Norge pågår det for tida forsking omkring potensialet til soppene Phomopsis cirsii og Ramularia cirsii mot dette ugraset.
Særlig åkertistel og den parasitterende rustsoppen Puccinia cirsii, har blitt studert nøye. Denne soppen kan bare formeres på levende planter og lar seg ikke dyrke på kunstig medium. Derfor er det bare konserveringsmetoden som eventuelt kan nyttes. Forskere i Nederland har studert effekten av å la flekkvis fordelte, sterkt rustinfiserte åkertistelbestander overvintre. Om våren vil disse bestandene raskt starte en ny epidemi. Infiserte åkertistler har brunrøde rustflekker under bladene, men siden skuddet overlever, ser de i utgangspunktet ikke ut til å la seg påvirke av rustsoppen. Rotsystemet til planten blir imidlertid latent infisert. Denne infeksjonen er systemisk, og rota vil neste vår sette enkelte unormalt høye, lysegrønne skudd som kjennetegnes av en søtlig duft. Disse plantene klarer ikke å sette nye utløpere, men de er kilde til ny smitte. Tilsvarende framgangsmåte studeres for rustsoppen P. lagenophora på åkersvineblom.
Mange sopper, men også bakterier, produserer giftstoffer, og noen av disse har ugrashemmende effekt. Slike mikrobielle stoffer kan brukes alene eller i kombinasjon med nytteorganismen for å øke effekten. Bladflekksoppen Ascochyta caulina vokser bare på meldestokk. Denne soppen produserer et plantehemmende giftstoff som hemmer meldestokk, men giften har virkning på mange tofrøblada arter hvis den brukes som et bio-herbicid. Selv om slike soppforbindelser er naturlig nedbrytbare, må de testes svært nøye med tanke på mulige helseskadelige effekter på mennesker og dyr.
Mikrobiologiske preparater på markedet innen biologisk ugrasbekjempelse
Det er foreløpig ingen mikrobiologiske preparater tilgjengelig på markedet i Norge. På verdensbasis er det bare noen få preparater som omsettes, som oftest for bruk i spesialkulturer.
I Nederland ble BioChon distribuert for kontroll av skuddannelse hos det amerikanske prydtreet romhegg, Prunus serotina, som har blitt en stor plage i Nederland. Midlet strykes på avkutta trestumper, og man har sett at 95 % av de nye skuddene dør innen to år. Hakatak og Stumpout er to tilsvarende midler som brukes mot innførte treslag i Sør-Afrika.
På appelsinplantasjer i Florida i USA har man oppnådd svært god bekjempelse av klatreplanten Morrenia odorata, som kveler sitrustrær, ved hjelp av preparatet DeVine, en formulering med soppen Phytophthora palmivora. I USA og Japan er preparater av bakteriene Xanthomonas campestris pv. poae og X. campestris pv. poa annua tilgjengelig for kontroll av tunrapp på golfbaner.
Fordeler med biologisk kontroll
Fordelene ved å bruke den klassiske biologiske kontrollmetoden er at nytteorganismene etter etablering vil reprodusere seg selv og spre seg til nye ugras uten ekstra kostnader. Dette gir lang virkningstid og blir relativt billig på lang sikt. Generelt for alle biologiske kontrollmetoder er at de ikke gir herbicidrester i miljøet. Det er også mindre fare for at ugraset blir resistent. Biologisk kontroll egner seg godt i situasjoner der en ugrasart dominerer og lager tette bestander som krever spesielle tiltak. Eksempler er åkertistel og burot. Andre ugras som egner seg for biologisk kontroll, er innførte arter, arter som opptrer i udyrket mark, eller har kun fjernt slektskap til våre kulturplanter.
Ulemper med biologisk kontroll
Klassisk biologisk kontroll er uaktuell så lenge metoden medfører introduksjon av en fremmed art som ikke er påvist her i landet. Et skrekkscenario ved bruk av biologisk kontroll er dårlige forundersøkelser som ikke avdekker at nytteorganismen skifter vertsplante. Dette var tilfellet i USA da man satte ut en eurasisk bille for bekjempelse av åkertistel. Dette endte med at nyttedyret gikk over til å bli en skadegjører ved at det skiftet preferanse til lokale tistelarter. Disse ble dermed kraftig redusert, og en allerede sårbar art av tistel ble nesten utryddet. En annen ulempe med klassisk biologisk kontroll er at metoden er treg i forhold til andre kontrolltiltak. I en amerikansk analyse regnet man med at det tok fem til femten år før man så målbar effekt på ugraset. Midlene til klassisk kontroll kan heller ikke lagres, og man er ikke garantert at alle de utsatte organismene overlever.
En begrensing for bruk av oversvømmelsesmetoden er at det vil være praktisk umulig å skaffe bioherbicider mot alle ugras i en åker. Biologisk bekjempelse av ugras er begrenset av nettopp den egenskapen som gjør en organisme nyttig, nemlig selektiviteten. Et ugrasbestand består som regel av mange arter, og når en art fjernes, blir den åpne plassen raskt overtatt av en annen art. En annen utfordring er at mangel på effekt i noen områder kan skyldes det lokale klimaet, slik at nytteorganismen ikke får etablert seg, eller at det finnes varianter av ugraset som er mer resistente enn normalt.
Vanskeligheter i produksjonen av smittestoff kan også stoppe nye mulige bioherbicider. Rustsoppene, som er effektive sjukdomsfremkallende organismer, kan ikke brukes som bioherbicid. De er så spesialiserte på verten (obligate parasitter) at ingen har lykkes med å framstille teliosporer, som kan infisere gjennom bladene, på kunstig medium. Smitten må derfor framskaffes ved å dyrke infiserte planter, noe som er dyrt og arealkrevende.
Integrering av biologisk kontroll med andre ugrasbekjempelsesmetoder
Som nevnt vil biologisk kontroll av ugras, sjelden kunne fungere som eneste tiltak.
I et ugrasbestand sammensatt av flere arter må biologisk kontroll som regel brukes i kombinasjon med mekaniske og dyrkingstekniske kontrolltiltak. Det er en forutsetning at biologien til ugraset og nytteorganismen er kjent, i tillegg til kjennskap til optimalt behandlingstidspunkt.
Hvorfor er det ingen biologiske ugrasmidler tilgjengelig på det norske markedet?
Biologisk bekjempelse av ugras er et relativt nytt forskningsområde her i landet. Midlene som er utviklet i USA og Australia er ikke aktuelle mot ugras som finnes i Norge, og med dyrkingsmetodene som nyttes her. Foreløpig har det vært utført et studium av soppen Ascochyta caulina mot den sommerettårige arten meldestokk. Soppen er her brukt som et bioherbicid som sprøytes ut på samme vis som de konvensjonelle ugrasmidlene etter oversvømmelsesmetoden. Klimaet i Norge om våren hemmer imidlertid infeksjon og etablering av nyttemikroben, og mye forskning gjenstår for å overvinne disse problemene (tekstboks 4.2).
Soppen Ascochyta caulina er en bladflekksjukdom på meldestokk og andre nært beslektede arter. Soppen ble testet for virkning mot meldestokk i forsøk utført i veksthus og på felt. Under optimale forhold i veksthus, det vil si 100 % fuktighet og 20 oC, forårsaket soppen kraftig angrep eller drepte ugraset. I feltforsøk med kålvekster på Ås, forårsaket soppen i en dose på 5 x 106 sporer per milliliter bare 0-26 % kontroll av meldestokken. Ved å kombinere nytteorganismen med 10 % anbefalt dose av Lentagran ble opp mot 88 % av ugraset kontrollert. Virkningen skilte seg ikke statistisk sikkert fra effekten av Lentagran alene. Grunnen til den dårlige virkningen av soppsporer alene skyldtes trolig ugunstige klimaforhold, som lav temperatur og lav luftfuktighet rett etter sprøyting. Dekking av det soppbehandla ugraset med fiberduk forbedret klimaforholdene noe, men ikke nok til at man så en klar økning i sjukdomsangrepet. For å øke væsketilførselen til soppsporene ble det derfor undersøkt om behandlinga av meldestokk kunne utføres rett før regn eller vanning av kulturen. Dessverre reduserte disse tiltakene mengda av soppsporer på bladoverflatene, slik at effekten på ugraset ble mindre. Det ble også undersøkt om strukturer som soppen produserer i flytende vekstmedium, kan tørkes og blandes i jord, hvor det er mindre fare for uttørking enn på bladoverflata. I innledende forsøk med meldestokkfrø, var effekten dårlig, men sjukdomsangrepet steg når dosen av soppstrukturer ble økt.
På alle planteoverflater, på og i jorda finnes en rekke ulike mikroorganismer, både sopp og bakterier, som utgjør en mikroflora. De fleste av disse skader ikke plantene, men lever av dødt vev eller av stoffer som plantene skiller ut. Mikroorganismene kan enten være upåvirket av hverandres nærvær, eller kan påvirkes positivt eller negativt av hverandre. Positiv påvirkning betyr for eksempel vekststimulering, mens negativ påvirkning kan være veksthemming eller induksjon av hvile (inaktivering). Negativ påvirkning kan oppstå ved konkurranse om plass og næring, eller ved at mikroorganismene skiller ut stoffer som hemmer veksten av andre arter. En slik form for konkurranse kalles antibiose. Et kjent eksempel er sopp i slekten Penicillium som skiller ut bakteriedrepende stoff, som brukes til behandling av ulike bakterieinfeksjoner hos mennesker og dyr. Det finnes også mikroorganismer som lever av å spise andre mikroorganismer. En mikroorganisme som lever av en planteskadegjører kalles en hyperparasitt («parasitt på parasitten»).
Denne naturlige formen for konkurranse mellom mikroorganismer ønsker vi å gjøre bruk av i bekjempelse av plantesjukdommer. Det er gitt mange definisjoner på biologisk bekjempelse opp gjennom årene. I denne spesifikke sammenhengen kan vi si at biologisk bekjempelse er å bruke levende mikroorganismer til å bekjempe mikroorganismer som forårsaker sjukdommer på planter. Slike nyttige mikroorganismer kalles for antagonister.
Ofte vil den naturlige mikrofloraen til plantene i stor grad beskytte mot angrep av sjukdomsorganismer. Det er vist at planter som er behandla med kjemiske plantevernmidler slik at mikrofloraen er forstyrret, kan bli verre rammet av sjukdom enn planter med uforstyrret mikroflora på overflaten. Ved epidemier vil den naturlige mikrofloraen imidlertid ikke gi tilstrekkelig beskyttelse. Plantevernpreparater bestående av antagonistiske mikroorganismer kan derfor bli et viktig hjelpemiddel i økologisk planteproduksjon. De kan tenkes brukt forebyggende, men også som direkte tiltak i tilfeller hvor en har et bestemt sjukdomsproblem i plantebestanden.
Tidlig på 1900-tallet ble det oppdaget at mikrofloraen i visse jordtyper var i stand til å forhindre sjukdom forårsaket av jordboende plantepatogener. Omkring 1930 fant man at den jordboende soppen Trichoderma lignorum reduserte skade på frøplanter forårsaket av plantepatogenet Rhizoctonia solani. Trichoderma-arter har siden vært blant de mest utprøvde soppene til biologisk bekjempelse av plantesjukdommer. Arter i denne slekten har antagonistisk effekt på mange ulike plantepatogener i blant andre soppslektene Pythium, Rhizoctonia og Botrytis.
Siden 1970-årene har forskningen på biologisk bekjempelse skutt fart som følge av økt kunnskap om skadelige sideeffekter av kjemiske plantevernmidler og økende restriksjoner på bruk av disse. I dag forskes det på antagonister, både sopp og bakterier, mot en rekke plantesjukdommer på alle kontinenter. I Norge har det vært gjort forskning på Trichoderma til bekjempelse av gråskimmel i eple og jordbær. I de senere årene har forskningen hovedsakelig fokusert på gråskimmel i jordbær, og antagonister som tilhører andre slekter enn Trichoderma, testes også ut.
Veien fra isolering av en antagonist til et ferdig produkt som kan tilbys planteprodusentene, er lang og omstendelig (tekstboks 4.3).
Isolering
Antagonisten må på et eller annet vis isoleres fra omgivelsene, fra plantemateriale, jord eller fra andre kilder (å isolere vil si å separere soppen fra omgivelsene og rendyrke den på kunstig næringsmedium).
Testing
Etter isolering av potensielle antagonister, er neste skritt å teste de ulike isolatenes evne til å kontrollere det aktuelle plantepatogenet. Den første delen av testingen foregår i laboratoriet under kontrollerte forhold. Her vurderes isolatenes evne til antagonisme, og for eksempel deres evne til å vokse ved lave temperaturer. En vanlig brukt metode er å pode antagonist og patogen på samme petriskål med næringsmedium, og se om antagonisten er i stand til å forhindre at patogenet vokser. Når man så har valgt ut potensielle antagonister i laboratoriet, er det aktuelt å gå videre med veksthusforsøk og feltforsøk med de mest lovende isolatene. For å få et riktig bilde av antagonistens evne til å begrense sjukdom på plantene, må feltforsøk gjentas over flere år og på forskjellige steder med ulikt lokalklima. Resultatet av feltforsøk er ikke bare avhengig av antagonistens evne til å hemme skadegjøreren. Det er også viktig å finne en hensiktsmessig metode for tilføring av antagonisten til plantebestandet (utsprøyting i suspensjon, innblanding i jorda, beising av frø osv.), og å finne passelig konsentrasjon og rett tidspunkt for applisering.
Kunnskap om patogen og antagonist
For å kunne bruke nyttesoppen mest mulig effektivt er det viktig at alle faktorer er så optimale som mulig. Dette forutsetter gode kunnskaper både om antagonisten og det biologiske systemet antagonisten skal introduseres i. Det er viktig å forsikre seg om at antagonisten ikke kan opptre som plantepatogen. Når det gjelder Trichoderma-slekten, finnes det for eksempel isolater av arten T. harzianum som er patogene på sjampinjong, slik at man kan få seg en overraskelse om man setter ut uidentifisert Trichoderma med tanke på å hindre sjukdom. Videre er det nødvendig å kjenne til hvilke klimatiske forhold antagonisten trenger for å vokse, hvilke næringsstoffer den er i stand til å benytte seg av, og hvilken virkningsmekanisme som gjør at den hemmer vekst av skadegjøreren. I tillegg er det nødvendig med en grundig risikovurdering av antagonisten, slik at en kan utelukke skadelige sideeffekter på mennesker og andre levende organismer. God kunnskap om plantepatogenets biologi er også en forutsetning. Om målet med antagonisten er å forhindre et plantepatogen i å infisere vertsplanten, er det viktig å kjenne til hvordan patogenet infiserer (med sporer som spres i vind, med sopphyfer fra andre infiserte plantedeler osv.), hvor den infiserer (røtter, bladplate, småplanter osv.), og hvilke klimaforhold den krever for å infisere (temperatur, fritt vann osv.). Det er også viktig å kjenne til hva slags vekstforhold vertsplanten tilbyr, særlig mengde og type næring. Nyere forskning har dessuten vist at antagonister utsettes for ulike former for forsvar fra de plantepatogene mikroorganismene, slik at dette også må tas hensyn til.
Produktutvikling
For at dyrkerne skal kunne bruke kommersielle preparater basert på mikroorganismer til bekjempelse av plantesjukdommer, må en bedrift være interessert i å lage et produkt. Produktutvikling omfatter også mange trinn som krever grundige undersøkelser. For det første må en finne fram til en enkel og rimelig metode for masseproduksjon av nytteorganismen. Deretter er det nødvendig med en formulering som gir tilstrekkelig lagringsdyktighet for preparatet, som jo består av levende organismer. Det må dokumenteres at det formulerte preparatet ikke kan skade miljøet eller være giftig for dyr eller mennesker. Til slutt må det søkes om godkjenning fra myndighetene til å markedsføre og selge produktet.
Tabell 4.1 Biologiske preparater tillatt brukt mot plantesjukdommer i Norge (per nov. 2005)
| Handelsnavn | Virksom organisme | Preparattype |
| Cedomon | Pseudomonas chlororaphis | Beisemiddel sopp |
| Mycostop | Streptomyces griseoviridis | Soppmiddel |
| Rotstop | Phlebiopsis gigantea | Soppmiddel |
(kilde: Mattilsynet)
Framtidsperspektiver; begrensninger, muligheter, utfordringer
Forskning viser at det ligger et stort potensiale i å bruke mikroorganismer til bekjempelse av plantesjukdommer. Flere kommersielle produkter er blitt utviklet i utlandet, nesten utelukkende for bruk i områder med varmere klima enn hva vi har her i landet. Derfor er forskning i Norge nå rettet mot å finne norske nyttesoppisolater som er bedre tilpasset vårt klima. Flere nye, lovende norske isolater er nær beslektet med kjente nyttesopper som er under utvikling i utlandet, noe som forhåpentligvis vil forenkle godkjenningsprosessen i Norge. Soppisolater fra slektene Gliocladium, Epicoccum, Ulocladium, Aureobasidium, Cladosporium og Humicola er blant de som nå er under utprøving for biologisk bekjempelse av plantesjukdommer. Fremdeles mangler vi tilstrekkelig kunnskap om samspillet mellom antagonister og patogener under ulike forhold for å kunne bruke mikroorganismer til effektiv sjukdomsbekjempelse, men den økende forskningsinnsatsen på dette området forventes å gi svar på viktige spørsmål i nær framtid. Viktige forskningsområder omfatter hvordan forskjellige miljøfaktorer påvirker effekten av ulike antagonister, hvilke mekanismer de ulike antagonistene tar i bruk mot sjukdomsorganismene i ulike mikrohabitater, om kombinasjon av ulike antagonister gir forbedret effekt, samt om formuleringen kan bidra til å øke antagonistenes overlevelse og aktivitet. Forskning er også rettet mot avklaring av infeksjonsmekanismer, f.eks. hvordan og under hvilke forhold den patogene soppen gråskimmel forårsaker latent smitte av gråskimmel i jordbær. Slik informasjon er viktig både for biologisk bekjempelse av sjukdommen og for avklaring av nyttige kulturtiltak, som både konvensjonell og økologisk planteproduksjon vil dra nytte av.
Biologisk bekjempelse, spesielt når det gjelder skadedyrkontroll, har vært brukt med stort hell siden 1890-årene.
Sesongintroduksjonsmetoden er en spesialisert utgave av oversvømmelsesmetoden. Dette er en viktig metode som brukes for skadedyrkontroll i europeiske veksthus. De naturlige fiendene slippes ut for å gi en øyeblikkelig virkning, men også for å gi en virkning gjennom hele vekstperioden mot skadedyr som har mange og kortvarige generasjoner, som spinnmidd, bladlus, mellus og trips. En lang rekke naturlig fiender til slik bruk masseproduseres av europeiske firmaer og kan kjøpes til bruk i norske veksthus (figur 4.2).
Organismer for biologisk bekjempelse av skadedyr vil vi i fortsettelsen dele inn i nyttedyr (insekter, midd og nematoder) og nyttemikroorganismer (sopp, bakterier, virus, protozoer).
I biologisk bekjempelse sorterer predatorer, parasitoider og nematoder under det vi kaller nyttedyr. Nyttedyrene kan ernære seg av, eller gjennomføre hele livssyklus inne i et av skadedyrene. Skadedyret kalles i denne sammenhengen for et byttedyr eller et vertdyr.
Nyttige insekter og midd som benyttes i biologisk bekjempelse deles inn i to hovedgrupper, predatorer (rovdyr) og parasitter (snyltere). Disse to gruppene er svært forskjellige når det gjelder livssyklus, søking etter byttedyr og næringsopptak.
Predatorer
En predator kan karakteriseres på følgende måte:
Parasitter
Ofte kalles en insektparasitt for en «parasittoid» for å skille den fra en ekte parasitt. En ekte parasitt dreper ikke vertdyret. Eksempler på ekte parasitter blant insektene er lopper og lus. En parasittoid derimot dreper alltid vertdyret for å kunne gjennomføre sin egen livssyklus.
En parasittoid kan karakteriseres på følgende måte (blir kalt en parasitt i fortsettelsen):
Livssyklus til en parasitt kan beskrives ved et eksempel fra en bladlussnylteveps:
Nematodenes biologi
De mikroskopiske rundormene, nematodene, regnes blant jordas mest tallrike flercellete dyr, og er mest kjent som skadedyr på planter og dyr. De fleste jordboende nematodene er rundt 0,5 til 2 mm lange og kan ikke sees med det blotte øye. Mange nematodearter lever på eller i insekter i en viss periode i deres livssyklus, uten at de nødvendigvis skader insektet. Vi kan inndele de forskjellige typer samspill mellom insekter og nematoder på følgende måte:
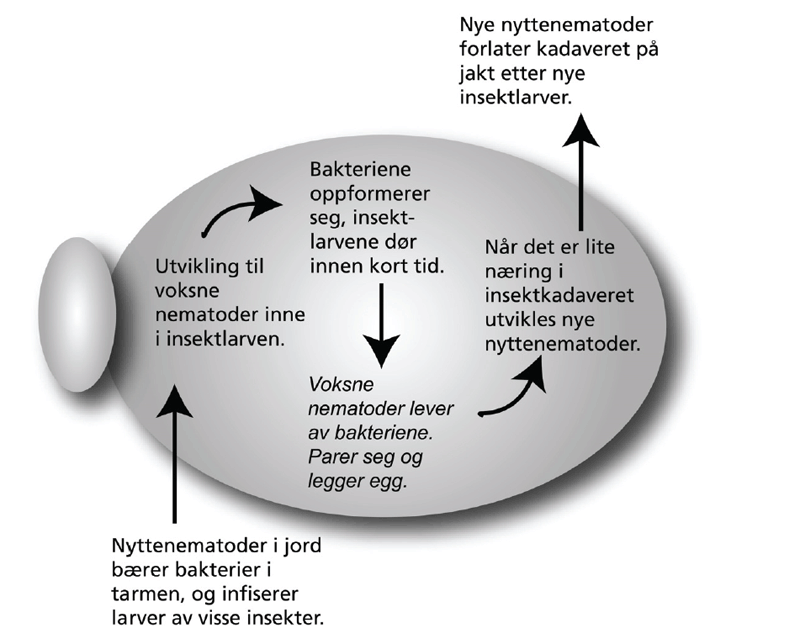
Nyttenematodene har et unikt forhold til en bakterie (Xenorhabdus eller Photorhabdus), der både bakterien og nematoden er avhengig av hverandre (figur 4.5). Ved å dyrke bakterien og nematoden i kunstig medium har man lykkes med å lage salgbare produkter av noen arter av nyttenematoder som kan brukes til biologisk bekjempelse av skadedyr i jord. Nyttenematoder lever av insektlarver (figur 4.5), men det stadiet av nyttenematodene som finnes utenfor insektsverten kan overvintre og overleve i flere måneder i jorda uten en vert. Nyttenematoder beveger seg lite i jord. Det kan være snakk om 5 til 25 cm, avhengig av art. Spredning foregår som oftest ved hjelp av insekter, dyr eller med jord. Vi vet forholdsvis lite om i hvilken grad nyttenematoder regulerer insektpopulasjoner i naturen, ettersom dette foreløpig er lite studert. Forskning på nyttenematodene har for det meste omhandlet utsetting ved hjelp av oversvømmelsesmetoden, praktisk bruk og produksjon.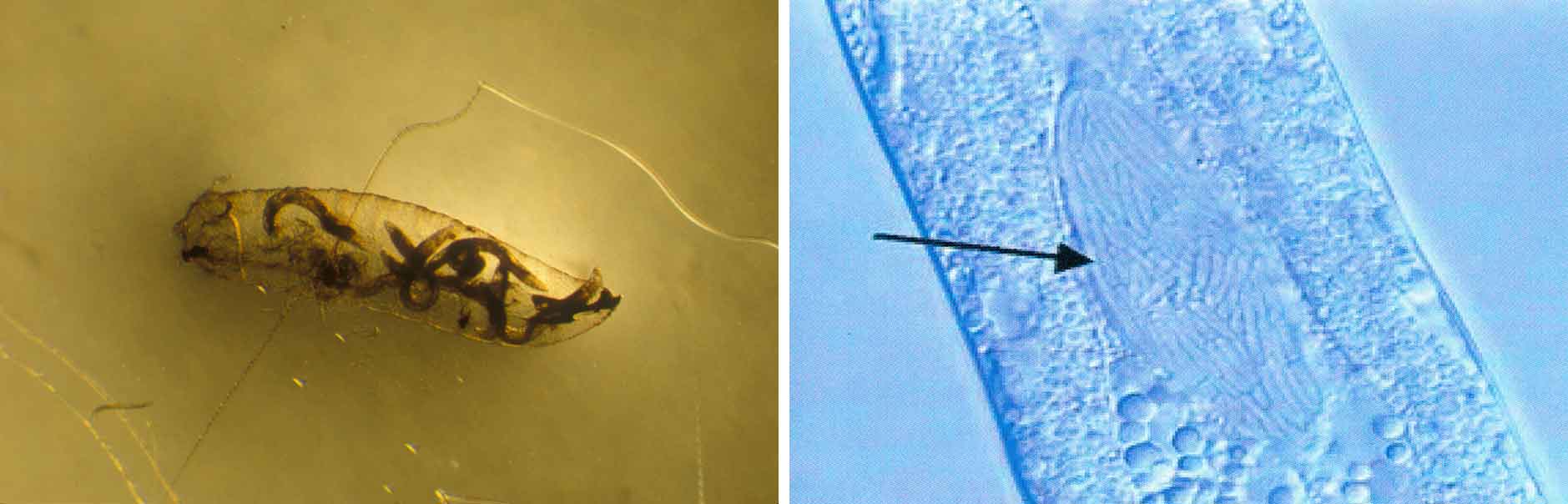
Teger
I flere familier av teger finnes arter som utelukkende er predatorer og dermed viktige nyttedyr, bl.a. i norske frukthager. De lever av mange ulike insektarter som små larver av sommerfugler og sugere, bladlus og midd. I familien nebbteger finnes tre arter innen slekten Anthocoris som er viktige nyttedyr i norske frukthager. Den vanligste arten er vanlig nebbtege. I frukthager på Østlandet finnes også de mindre dvergnebbtege (figur 4.7) som nyttedyr. Blant bladtegene finnes mange viktige skadedyr på planter. Men noen arter av bladteger er nyttedyr, da de helt eller delvis lever som predatorer, særlig av bladlus og midd. Eksempler på nyttige bladteger i frukthager er svartknetege og grønnspraglet langfottege.
Nettvinger
Nettvingene er lette å kjenne igjen på det tette ribbenettet i vingene og de lange antennene. Larvene er grådige predatorer som griper byttedyret med de kraftige fremadrettete kjevene og suger ut innholdet. Vi finner viktige nyttedyr i følgende tre familier: gulløyer (figur 4.6), bladlusløver og middløver. Larver av gulløyer spiser vesentlig midd, bladlus og små sommerfugllarver. Noen arter av gulløyer er også predatorer som voksne. Hos bladlusløver lever både de voksne insektene og larvene av bladlus og midd. Larvene av gulløyer og bladlusløver blir opptil 7-8 mm lange, mens de fullvoksne larvene av middløvene er små, 2-3 mm lange. Larvene av middløver har betydning som nyttedyr i frukthager, der de lever av frukttremidd og andre midd.
Tovinger
Gallmygg er først og fremst kjent som skadedyr på en del kulturvekster, men noen få arter lever som predatorer på midd og insekter. I bladluskolonier kan en finne de små rødlige fotløse larvene til bladlusgallmyggen.
Det finnes nærmere 300 norske arter av blomsterfluer. De fleste har svart- og gulstripet bakkropp og kan minne om stikkeveps. Larvene til blomsterfluer er glupske predatorer på bladlus. De voksne fluene legger eggene sine i bladluskolonier. Larvene er flate, fotløse og med et redusert hode, men med kraftige kjever. De har torner og pigger på leddene og er vanligvis hvite, oransje eller grønne med mørke fargetegninger (figur 4.6). En larve kan sette til livs flere hundre bladlus før den er fullvoksen.
Biller
Det er vel 50 arter av marihøner i Norge. To vanlige arter, 7-prikket marihøne og 2-prikket marihøne (figur 4.6) er begge røde med svarte prikker, mens andre arter kan for eksempel være gule med svarte tegninger. Larvene er gråblå med gule flekker og med små utvekster på leddene. Marihønene er med få unntak predatorer både som voksne og larver. De lever hovedsakelig av bladlus, men noen få arter spiser hovedsakelig midd. I år med sterke bladlusangrep kan særlig 7-prikket marihøne bli svært tallrik.
Det finnes ca. 900 norske arter av kortvinger. Både voksne biller og larvene er predatorer, og mange arter er viktige nyttedyr i norske åkre. De fleste kortvingene er 2–15 mm lange, slanke og er mørke som voksne. De korte dekkvingene som ikke dekker hele bakkroppen, er karakteristisk for kortvingene (figur 4.7). Mange arter er nattaktive.
Edderkoppdyr
Midd er en artsrik gruppe edderkoppdyr med små arter, de fleste under 0,5 mm lange. Hos midd er for- og bakkroppen sammensmeltet og kroppen er uten synlig leddeling. Midd har 4 par bein, med unntak av det første stadiet som klekker fra egget (larve) som har 3 par bein. Mange arter suger på planter og er viktige skadedyr, men vi finner også mange viktige predatorer blant middene innen familien rovmidd (figur 4.7). Disse lever av plantesugende midd og små insekter som trips m.fl.
Det er kjent ca. 500 arter av edderkopper i Norge. De fleste edderkoppene er 3-15 mm lange og mørke. De er utelukkende rovdyr. I en undersøkelse av norske åkre ble det funnet ca. 80 arter. Mattevevere (særlig Oedothorax apicatus, Erigone atra og E. dentipalpis) og ulveedderkopper (særlig Pardosa palustris) var de absolutt vanligste. Aktiviteten til disse edderkoppene var størst i juni.
De insektparasittene som brukes i biologisk bekjempelse tilhører to insektordener, tovinger og årevinger. Kun én familie av tovinger er aktuell, nemlig snyltefluer. De viktigste parasittene er årevingene.
Snyltefluer
Det finnes ca. 200 arter av snyltefluer i Norge. De fleste artene parasitterer sommerfugllarver. Betydning av snyltefluer som naturlige fiender er lite undersøkt i Norge.
Snylteveps
Snyltevepsene tilhører underordenen stilkveps og har en tydelig «vepsetalje», dvs. en tydelig innsnevret del foran på bakkroppen. Snyltevepsene inndeles i flere overfamilier med en lang rekke familier innen hver gruppe. Artsantallet i Norge er dårlig undersøkt. Det er 728 kjente arter per i dag, men trolig finnes det ca. 7000 arter av snylteveps i Norge. Biologien er også svært dårlig kjent. Her skal vi bare omtale noen få viktige familier av snylteveps.
Ichneumonoidea
Overfamilien Ichneumonoidea inneholder mange viktige parasitter, bl.a. i familiene ichneumonider, braconider og bladlussnylteveps. Ichneumonidene er relativt store arter av snylteveps som parasitterer larvene til en lang rekke arter av sommerfugler. Braconidene er ofte mindre arter enn ichneumonidene. De parasitterer mange forskjellige insekter, men også her er sommerfugllarver viktige vertdyr. Velkjent er Cotesia glomerata som parasitterer stor kålsommerfugl. Bladlussnylteveps har en meget enhetlig biologi, da alle artene utelukkende parasitterer bladlus. Disse snyltevepsene er små, 2-3 mm. Livssyklusen er beskrevet tidligere. Vanlige slekter er Aphidius (gir gråbrune mumier) og Ephedrus (gir svarte mumier). Artene i slekten Praon gir lyse «dobbeltmumier», der bladsnyltevepsens puppekokong blir liggende under selve det tomme bladlusskallet (figur 4.6).
Chalcidoidea (chalcider)
Dette er en overfamilie med små arter av snylteveps, 2-3 mm lange, ofte med vakre blå og grønne metallglinsende farger. De er lette å kjenne igjen på fargene, det reduserte ribbenettet i vingene og de knebøyde og klubbeformete antennene. Mange arter er hyperparasitter, dvs. at de er parasitter på insekter som allerede er parasittert av en annen art. Hyperparasitter kan derfor være et negativt innslag i biologisk bekjempelse av skadedyr. I familien Aphilinidae finner vi arten Encarsia formosa som brukes i norske veksthus for å bekjempe veksthusmellus. Ørsmå snylteveps, 0,5-1,5 mm, i familien Trichogrammatidae har spesialisert seg på utelukkende å parasittere og gjennomføre livssyklus inne i et enkelt insektegg.
Naturlig forekomst av nyttenematoder i Norge
Nyttenematoder forekommer naturlig i jord og har blitt påvist i alle verdensdeler med unntak av Antarktis. I Norge er det påvist flere arter i jordbruks- og skogsjord fra hele landet. I flere undersøkelser er det påvist flest arter i slekten Steinernema bl.a. S. feltiae, S. kraussei og S. carpocapsae. Heterorhabditis-slekten er ikke like vanlig forekommende som Steinernema, men to arter er påvist så langt, H. megidis og en uidentifisert art. De nevnte artene er aktuelle i biologisk bekjempelse både i veksthus og på friland. Kunnskap om utbredelse av nyttenematoder er basert på kartlegging og analysering av jordprøver. Det er sjeldent en observerer døde nematodeinfiserte insektlarver i naturen fordi de går fort i oppløsning. Figur 4.8 viser hageoldenborrelarver naturlig infisert med en nyttenematode (Heterorhabditis megidis) i en plen i Sør-Norge.
Per nov. 2005 er 15 ulike predator- og parasittarter og 3 ulike nematodearter godkjent for biologisk bekjempelse i Norge. Se tabell 4.2 for oversikt over aktuelle nyttedyr som er å få kjøpt.
Tabell 4.2 Nyttedyr som selges på det norske markedet per i dag 1)
| Nyttedyr | Virkeområde | Bruksområde |
| Predatorer: | ||
| Amblyseius cucumeris | Rovmidd mot tripsegg og -nymfer, samt dvergmidd og spinnmidd | Plantekulturer i veksthus |
| Aphidoletes aphidimyza | Gallmygg (larver) mot bladlus | Plantekulturer i veksthus |
| Cryptolaemus montrouzieri | Marihøner mot ullus (Planococcus og Pseudococcus) | Plantekulturer (først og fremst prydplanter) i veksthus (ikke tomat) |
| Feltiella acarisuga | Gallmygg mot egg, larver, nymfer og voksne spinnmidd (inkl. dvalehunner) | Plantekulturer i veksthus |
| Hypoaspis miles | Rovmidd mot jordboende stadier av hærmygg (vannfluer, trips og spretthaler) | Plantekulturer i veksthus og ved produksjon av matsopp |
| H. aculeifer | Rovmidd mot jordboende stadier av hærmygg, løkmidd (vannfluer, trips og spretthaler) | Plantekulturer i veksthus og ved produksjon av matsopp |
| Macrolophus caliginosus | Rovtege mot veksthus- og bomullsmellus (spinnmidd, bladlus, trips, sommerfuglegg og blad-minerfluelarver) | Plantekulturer i veksthus |
| Orius majusculus | Rovtege mot nymfer og voksne trips | Plantekulturer i veksthus |
| Phytoseiulus persimilis | Rovmidd mot egg, larver, nymfer og voksne spinnmidd | Plantekulturer i veksthus og tunnel |
| Parasitoider: | ||
| Aphidius colemani | Snylteveps mot små bladlus som ferskenbladlus og agurkbladlus | Plantekulturer i veksthus |
| A. ervi | Snylteveps mot potetbladlus, grønnflekket veksthusbladlus, rosebladlus m.fl. | Plantekulturer i veksthus |
| Dacnusa sibirica | Snylteveps mot minerfluelarver i Liriomyza - og Chromatomyia –slekten | Plantekulturer i veksthus |
| Diglyphus isaea | Snylteveps mot minerfluelarver i Liriomyza - og Chromatomyia –slekten | Plantekulturer i veksthus |
| Encarsia formosa | Snylteveps mot veksthus- og bomullsmellus Plantekulturer i veksthus | |
| Eretmocerus eremicus | Snylteveps mot veksthus- og bomullsmellus | Plantekulturer i veksthus |
| Nematoder: | ||
| Steinernema feltiae | Hærmygg, trips | Prydplanter (mest veksthus), hobby |
| S. kraussei | Rotsnutebiller2) | Prydplanter og jordbær (friland, veksthus, planteskoler, hobby) |
| Heterorhabditis megidis | Rotsnutebiller3) og hageoldenborre | Prydplanter og jordbær (friland, veksthus, planteskoler, grøntarealer, golfbaner, hobby) |
1) NB! Bak hvert nyttedyr kan det skjule seg flere ulike produkter som er effektive mot ulike skadedyr.
2) Bl.a. veksthussnutebille for behandling ved lav jordtemperatur (6-12 oC)
3) Bl.a. veksthussnutebille for behandling ved høy jordtemperatur (12-25 oC)
Veksthus
Utsetting av predatorer og parasitter ved hjelp av oversvømmelsesmetoden er først og fremst brukt i veksthus i Norge. Predatorer og parasitter kan gi meget god bekjempelse av skadedyr i veksthus når de brukes forebyggende, eller hvis de slippes ut med en gang angrep av skadedyr oppstår. Ved sterke angrep blir bekjempelsen ofte utilstrekkelig fordi nyttedyrene ikke klarer å bekjempe skadedyrene før plantene får for stor skade. Kontinuerlig overvåkning av skadedyrsituasjonen er derfor viktig. Utslippsmengde og hyppighet vil variere med angrepsgrad, kultur og produksjonsområde. Er du i tvil om valg av nytteorganisme eller utslippsmengde er det derfor lurt å kontakte veiledningstjenesten. Nyttedyrene må bestilles i god tid før de skal brukes fordi det tar noen dager å få dem tilsendt. Importører og forhandlere av nytteorganismer er blant andre L.O.G., Gartnersenteret Lier, Fruktlageret Handel AS og VekstMiljø AS. Nyttedyrene bør helst brukes med en gang de mottas. Dersom de må lagres, er det imidlertid viktig å skaffe seg rede på hvordan dette bør gjøres. Dersom de lagres for varmt, kan nyttedyrene lett dø, og du vil ikke få noen effekt av utslippet. De fleste nyttedyr kan ikke lagres mer enn i 1-2 døgn, selv om de lagres under optimale forhold.
Friland
På friland er svært få predatorer og parasitter godkjent for utsetting, men konserveringsmetoden, som legger til rette for best mulig utnytting av predatorer og parasitter som forekommer naturlig i et felt, er aktuell. For å kunne bruke konserveringsmetoden er det nødvendig å ha gode kunnskaper om predatorenes og parasittenes biologi og levevis slik at en kan si noe om under hvilke forhold de fungerer mest effektivt. Flere norske studier har derfor dreiet seg om biologi og levevis hos predatorer og parasitter.
Blant annet har en funnet at vanlig tetthet av løpebiller i norske åkre om sommeren er 5–10 individer per m2, og at mange arter senhøstes trekker inn til åkerkantene, hvor de samler seg for overvintring. Åkerkanter er altså viktig for å bedre overlevelsen av løpebiller. En kan også dele opp større åkre ved å anlegge 2 m brede grasstriper. Både larvene og de voksne billene er predatorer. De kan ha betydning som naturlige fiender for flere skadedyr i jordbruket, for eksempel bladlus i korn og kålfluelarver i kålvekster. I norske åkre er det også funnet ca. 230 arter av kortvinger, men bare omkring 20 arter er dominerende. Det forekommer vanligvis 10-80 individer per m2. De fleste kortvingene overvintrer som voksne i åkerkantene, og her er det vinterstid funnet ca. 200 individer per m2. Larvene finnes i jorda om sommeren. Biologien til kortvingene er ikke like godt undersøkt som hos løpebillene, men mange arter har stor betydning som predatorer på egg og larver av mindre skadedyr i grønnsaker og jordbruksvekster. De viktigste predatorene antas å tilhøre slektene Philonthus og Tachyporus.
Rollen til edderkoppene som nyttedyr i jordbruket er lite undersøkt, men utenlandske undersøkelser viser at de kan ha betydning som predatorer på skadedyr, særlig på forsommeren. I et norsk pilotstudium hvor forekomsten av edderkopper i en økologisk og en konvensjonell jordbæråker har blitt undersøkt, fant en flere edderkopparter, og dessuten flere individer av unge (juvenile) edderkopper i det økologiske enn i det konvensjonelle jordbærfeltet. Rovmidd har vist seg å ha størst betydning som viktige nyttedyr i frukthager på friland i Norge. På de fem viktigste fruktslagene i Norge er det funnet 25 arter av rovmidd. Fem av disse kan forekomme svært tallrikt og er effektive predatorer på mange plantespisende midd, særlig på frukttremidd. Gjennom en vekstsesong klarer de å redusere sterke angrep av frukttremidd til et minimum. Det foregår forsøk også med biologisk bekjempelse av jordbærmidd med bruk av rovmidden Neoseiulus (Amblyseius) cucumeris i jordbær på friland (oversvømmelsesmetoden) i Norge, men metoden er enda ikke godkjent for praktisk bruk her til lands. N. cucumeris er utbredt langt nordover i Europa (Tyskland, England og Nederland), men det er knyttet stor usikkerhet til hvorvidt arten finnes naturlig i Norge. N. cucumeris brukes imidlertid til kontroll av jordbærmidd på friland i Danmark, Finland og Sverige.
Praktisk bruk av nematoder i Norge
I Norge brukes nematoder mot skadedyr både i veksthus og på friland. For eksempel brukes nyttenematoder (Steinernema feltiae) til bekjempelse av hærmygg i veksthus. Andre områder der nematoder brukes i Norge er mot rotsnutebiller i jordbær og i planteskoler. Alle nematodepreparatene kan også brukes som hobbypreparater. Nematoder brukes i hovedsak mot jordboende insektlarver, men nylig er det også vist at nematodene kan brukes på bladverk i veksthus mot trips. Nematodeartene som selges på det norske markedet er påvist naturlig forekommende i landet.
Nematodeproduktene er formulert slik at det kan oppbevares kjølig i noen få måneder, men det er anbefalt å bestille produktene like før behandling for å få best mulig kvalitet. Produktene skal løses opp i vann og deretter vannes på området som skal behandles. Utvanning skal foregå på det tidspunkt skadedyrene er mottakelige. Det er ofte snakk om en dose på 500 000 nematoder per effektiv m2 (oversvømmelsesmetoden). Det pågår forskning for å kunne redusere denne høye dosen, og foreløpige resultater antyder at flere utvanninger med en lav dose er gunstigere enn en tilføring av en høy dose. Andre forsøk har vist at dypping av småplanter i en nematodeløsning før utplanting kan være effektivt. Bruk av nyttenematoder gjennom dryppvanningsanlegg er også mulig, men lite utprøvd. Det pågår internasjonal og nasjonal forskning for å forbedre bruken av nyttenematoder ute i felt og inne i veksthus. Det er vist få negative effekter ved bruk av nematoder, men utslipp kan gi en lokal negativ effekt på nytteinsekter som for eksempel larver av løpebiller kort tid etter nematodeutslipp. Bruk av nyttenematoder egner seg svært godt i integrert bekjempelse av insekter, der flere miljøvennlige metoder tas i bruk for å redusere skade gjort av insekter.
Innen fagfeltet biologisk kontroll av skadedyr legges det stor vekt på nyttedyr som naturlige reguleringsfaktorer og effektive biologiske kontrollagenter for skadedyr. Naturlig forekommende mikroorganismer som sopp, bakterier og protozoer samt virus utgjør imidlertid også en viktig dødelighetsfaktor for mange skadedyr. Funn gjort både i Norge og utlandet peker i retning av at nyttemikroorganismer bør få en viktigere plass i biologisk kontroll av skadedyr.
Soppenes biologi
Soppene har en unik stilling blant insekt- og middsjukdommene (patogenene) fordi de infiserer ved å trenge direkte gjennom skadedyrets kutikula («hud»). Skadedyret behøver derfor ikke å spise soppen for å bli infisert, og dermed blir også sugende skadedyr lett infisert av sopp. Det finnes rundt 750 forskjellige sopparter som infiserer insekter og midd. De fleste av disse soppene hører til klasse Hyphomycetes eller til orden Entomophthorales.
Blant hyphomycetene finner vi sopparter som lett kan dyrkes på kunstige medier, og dermed også masseoppformeres. Sopp som tilhører denne klassen, er ofte bredspektra, og kan dermed drepe insekt- og middarter som står forholdsvis langt fra hverandre systematisk. En naturlig epidemisk utvikling av disse soppene oppstår vanligvis bare i jordlevende insekt- og middarter. De fleste mikrobiologiske plantevernproduktene er basert på sopp fra klasse Hyphomycetes.
Insekt- og middpatogene sopper innen orden Entomophthorales er hyphomycetenes rake motsetning. De er vanskelige å dyrke på kunstige medier, men er kjent for å forårsake spektakulære epidemier på insekter og midd som lever på bladverk og overjordiske plantedeler. Disse soppene er ikke bredspektra, men har et smalt vertsregister. Insektpatogene sopper innen Entomophthorales er særdeles vanskelige å masseoppformere og finnes ikke som ferdig formulerte plantevernprodukter. Entomophthorales kan likevel brukes i biologisk bekjempelse ved at dyrkerne legger forholdene best mulig til rette for soppen (konserveringsmetoden). Det finnes flere vellykkede eksempler på dette fra utlandet.
Bakterienes biologi
Bakterier infiserer for det meste gjennom munnen, og må derfor spises. I mikrobiologisk kontroll av skadedyr har det spesielt blitt fokusert på bakterier innen familien Bacilliceae og da spesielt på artene Bacillus thuringiensis (Bt) og B. popilliae (tekstboks 4.4).
Bacillus thuringiensis (Bt) er en bakterie som danner sporer (kalt endosporer) inne i et sporehus. Når sporene produseres inne i sporehuset, blir det overproduksjon av proteiner. Disse proteinene danner krystaller inne i sporehuset, men utenfor sporene. De tilhører en gruppe proteiner som er giftige (toksiske) overfor ulike insektgrupper, og de kalles endotoksiner. Figur 4.9 viser virkningsmekanismen for Bt i en sommerfugllarve. 1) For at Bt skal virke må insektet først spise Bt-sporehuset som inneholder sporer og krystaller. 2) Krystallet løser seg deretter opp i magen til vertdyret, og toksinene blir aktivert. For at krystallet skal kunne løse seg opp og toksinene aktiveres, må det være spesielt høy pH i magesekken til vertdyret. 3) Når toksinet er aktivert, binder det seg til spesielle steder på magesekkcellene til vertdyret, den osmotiske balansen forstyrres, cellene sveller og ødelegges, og insektet dør etter kort tid. 4) Når cellene ødelegges kommer insektblodet inn i magesekken. Dette fører til forhold (lav pH og mye næringsstoffer) som får Bt-sporen til å spire og danne vegetative celler som formerer seg inne i insektet. Infeksjonssyklusen er avsluttet når Bt har brutt ned insektet og Bt ender opp i miljøet som sporer. For toksiner i Bt-planter er virkningsmekanismen mindre spesifikk enn for et Bt-sprøytemiddel fordi det ikke trengs en aktivering av noe krystall. Bt-planter er dermed også mindre vertsspesifikke enn Bt-sprøytemidler.
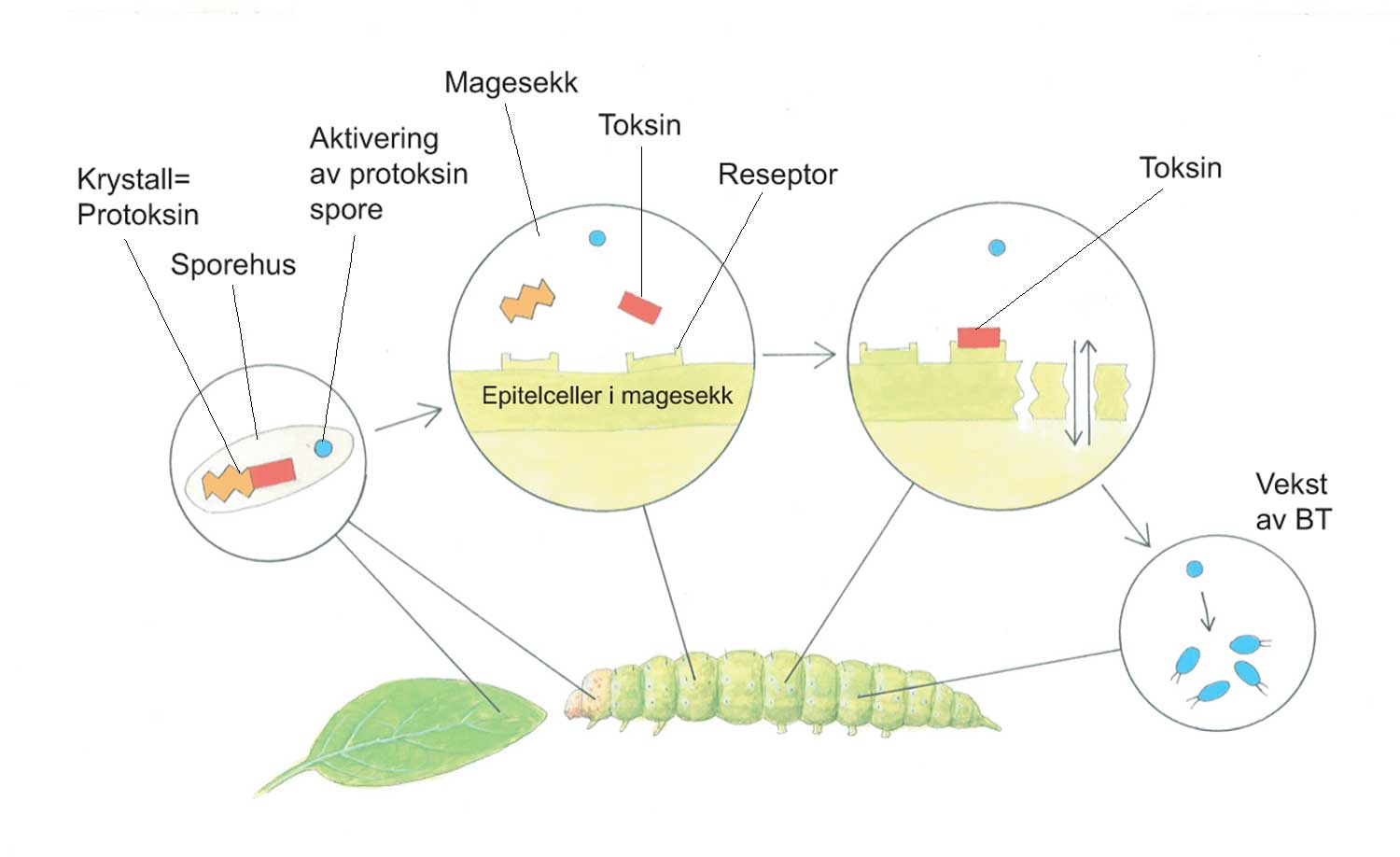
Soppene
Soppene er den gruppen nyttemikroorganismer som foreløpig er studert mest i Norge, og det er gjort undersøkelser av sopp som naturlig dødelighetsfaktor for mange viktige skadedyr. Noen av disse undersøkelsene presenteres nedenfor.
Kirsebærbladlus er et årvisst skadedyr i kirsebær (figur 4.10). I utsprøyta kirsebærfelt har en funnet flere forskjellige sopparter som dreper kirsebærbladlus. Et betydelig antall bladlus blir infisert og drept av disse soppene, men infeksjonsprosenten varierer sterkt gjennom sesongen. Også snylteveps er en dødelighetsfaktor for kirsebærbladlus. Snyltevepsene er det flest av tidlig i sesongen. Senere dominerer nyttesoppene. Rød eplebladlus og grønn eplebladlus har også vist seg å ha fiender blant flere av de samme soppene som infiserer og dreper kirsebærbladlusa.
Neozygites floridana er en sopp som infiserer og dreper veksthusspinnmidd (figur 4.10). Veksthusspinnmidd kan være et plagsomt skadedyr i jordbær, spesielt under tørre og varme forhold, og når det sprøytes mye med kjemiske plantevernmidler mot andre skadegjørere. Studier fra USA viser at N. floridana er en av de viktigste dødelighetsfaktorene for veksthusspinnmidd i mais og soyabønne. Funnene har ført til at det i USA er utviklet integrerte dyrkingssystemer som er med på å legge forholdene til rette for denne nyttesoppen i mais og soya. I Norge er N. floridana funnet på veksthusspinnmidd i jordbær, og det undersøkes om den er viktig for kontrollen av veksthusspinnmidd i frilandsjordbær.
Soppartene Entomophthora muscae (figur 4.10) og Strongwellsea castrans infiserer og dreper voksne individer av stor og liten kålflue i Norge. Stor og liten kålflue er begge alvorlige skadedyr i kålvekster, og larvene til disse fluene angriper røttene til kålplanter. I en undersøkelse gjort i et usprøytet hodekålfelt, ble det funnet at opptil 48 % av de voksne kålfluene ble infisert og drept av E. muscae i løpet av en sesong, og opptil 18 % kunne dø med S. castrans tegn. Dette tyder på at disse soppartene kan være en viktig dødelighetsfaktor for kålfluene, og at de derfor kan ha et potensiale for kontroll av kålfluene (tekstboks 4.5).
Både Entomophthora muscae og Strongwellsea castrans har fascinerende livssykluser og utspekulerte strategier for spredning og infeksjon. En kålflue som er infisert av E. muscae dør etter omtrent 7 dager. I begynnelsen av infeksjonen oppfører den infiserte flua seg tilsynelatende normalt, men lenger ut i infeksjonsforløpet vil flua bli mer desorientert, og etter hvert vil den plassere seg høyt i vegetasjonen med bakparten i været. I denne posisjonen vil flua dø, og soppen selv sørger for at flua blir skikkelig festet til planten den har plassert seg på. Dette gjøres ved at soppen produserer festeanordninger (rhizoider) som vokser ut gjennom fluas munndeler. Med flua festet på denne måten trenger soppens hvite sporebærere seg ut gjennom de tynnhudede stedene på fluas bakkropp, og sporene blir aktivt skutt ut for så å infisere andre fluer. Døde sporulerende hunner som står på denne måten med bakparten i været, blir ofte forsøkt paret av paringsklare hanner, og smitten kan dermed lett overføres til friske hunner som blir paret av de samme hannene.
Spredningsstrategien til S. castrans er også forunderlig. En flue infisert med denne soppen kan leve i rundt to uker. En frisk flue lever i maksimalt fem, og en infisert hunnflue kunne derfor ha fått tid til å legge en del egg før den dør. Navnet «castrans» antyder imidlertid at dette ikke er tilfelle. Kålfluer angrepet av S. castrans utvikler et stort hull, som kan sees med det blotte øye, på undersiden av bakkroppen. Fra dette hullet blir sporene skutt ut, og fluene kan leve i rundt en uke etter at hullet har utviklet seg. Hullet forårsaker ikke større atferdsendringer hos flua, og den kan dermed fly rundt som vanlig, samtidig som den skyter ut sine dødbringende sporer som infiserer andre kålfluer.
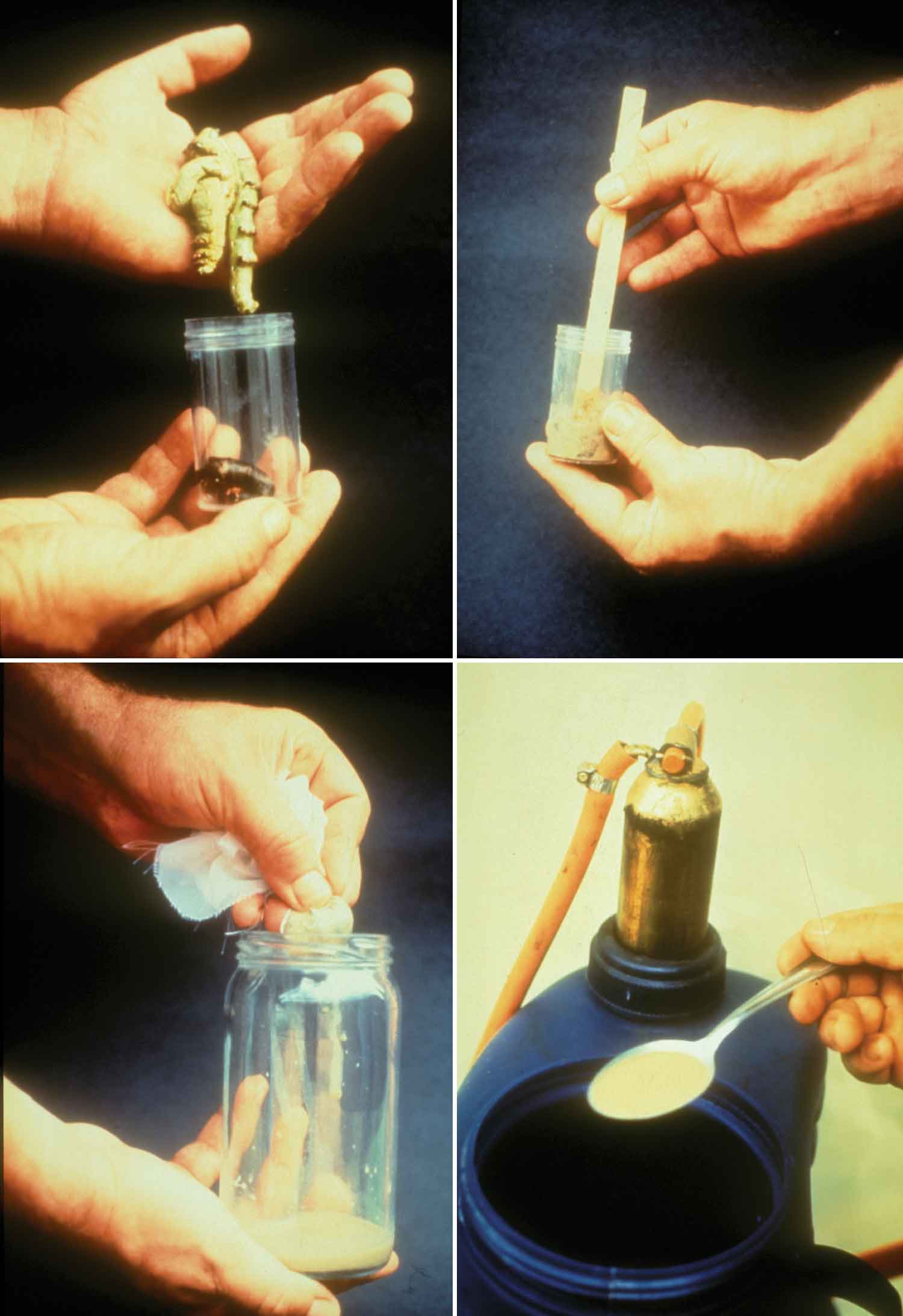
Et par gode eksempler på lokal oppformering og bruk av virus har en fra bekjempelsen av sommerfugllarvene Erinnys ello og Anticarsia gemmatalis i Brasil. Begge skadedyrene blir bekjempet ved at en samler inn virusinfiserte larver en sesong og oppbevarer dem i fryseren til påfølgende sesong. Deretter lages det et viruspreparat som sprøytes på nyklekte larver. E. ello bekjempes ved hjelp av et granulovirus (GV), og A. gemmatalis av et nucleopolyhedrovirus (AgNPV). Et ferdig formulert produkt av AgNPV er også produsert, og dette preparatet brukes nå på ca. én million hektar årlig i Brasil.
Dersom en ønsker å utnytte nyttemikroorganismer til kontroll av skadedyr, benytter en seg av tilsvarende strategier som for annen biologisk kontroll av skadedyr.
Vil en utvikle en strategi som legger til rette for konservering og fremming av nyttemikrobene, er det viktig å ha gode kunnskaper om nyttemikrobenes biologi og livssyklus. Vet en hvordan nyttemikrobene overvintrer, og under hvilke forhold de spres og trives, er det også lettere å vite hvordan man kan bevare og fremme dem. Både vanning, alternative vertdyr og kantvegetasjon har vist seg å spille en viktig rolle for spredning av nyttemikroorganismer som dreper insekter og midd. Mange av soppene som dreper skadedyr, blir negativt påvirket av plantevernmidler, spesielt kjemiske soppmidler. Av godkjente plantevernmidler på DEBIOs liste som muligens kunne være uheldig for nyttesoppene, finner vi svovelpreparatene. Forsøk gjort i utlandet viser imidlertid at svovel kan ha, men har vanligvis ikke noen negativ effekt på nyttesoppene.
Metodene som er nevnt ovenfor, representerer en forsiktig påvirkning av patogen, skadedyr og miljø. Den metoden som brukes i størst omfang i dag er imidlertid nyttemikrobene brukt som biopesticid. Det finnes en rekke mikrobiologiske skadedyrmidler på verdensmarkedet, og stadig kommer nye til (tabell 4.3). Ferdig formulerte skadedyrmidler basert på mikroorganismer mot skadedyr på friland finnes foreløpig ikke i Norge. Det finnes imidlertid flere godkjente soppbaserte preparater til bruk mot skadedyr i veksthus.
Tabell 4.3 Noen av de viktigste nyttemikrobene som finnes i ferdig formulerte produkter på det internasjonale markedet 1)
| Patogen | Virkeområde | Bruksområde |
| Bakterier: | ||
| Bacillus popilliae | Japanbille (eng. cockchafer ), andre skarabider | |
| Bacillus thuringiensis **) | Sommerfugllarver, larver av hærmygg og biller | Grønnsaker og prydplanter på friland og i veksthus |
| Sopp: | ||
| Beauveria bassiana | Plantesugere, biller og sommerfugler | Bladverk og jord |
| Beauveria brogniartii | Oldenborrer, andre billearter, plantesugere, sommerfugler og tovinger | Eng, beite, plen, blomster, grønnsaker, poteter og oljepalmer |
| Metarhizium anisopliae | Biller i jord | Prydplanter i veksthus og på friland |
| Nomuraea rileyi | Sommerfugler | Prydplanter, grønnsaker, poteter, oljepalmer |
| Paecilomyces fumosoroseus *) | Veksthusmellus, bomullsmellus, bladlus, trips, biller og spinnmidd | Veksthus, prydplanter, grønnsaker, frukt |
| Verticillium lecanii *) | Bladlus, mellus, trips, spinnmidd og tovinger | Grønnsaker og prydplanter i veksthus |
| Virus: | ||
| AcMNPV | Autographa californica (nattfly) | Luserne, bringebær, salat, bomull |
| OpMNPV | Orgyia pseudotsugata (sommerfugl/ Lymantriidae) | Bartrær |
| LdMNPV | Løvskognonne | Løvskog |
| SeMNPV | Spodoptera exigua (multiresistent sommerfugl) | Grønnsaker og prydplanter |
| HzSNPV | Helicoverpa zea (multiresistent sommerfugl) | Kikert, bomull, mais, okra, tomater, prydplanter m.m. |
| NsSNPV | Rød furubarveps | Furuskog |
| AoGV | Fruktskallvikler | Eplehager |
| CpGV | Eplevikler | Eplehager |
1) NB! Bak hvert patogen kan det skjule seg flere ulike isolat og flere ulike produkter som er effektive
mot ulike skadedyr. *) Godkjent i Norge. **) Søkt godkjent i Norge
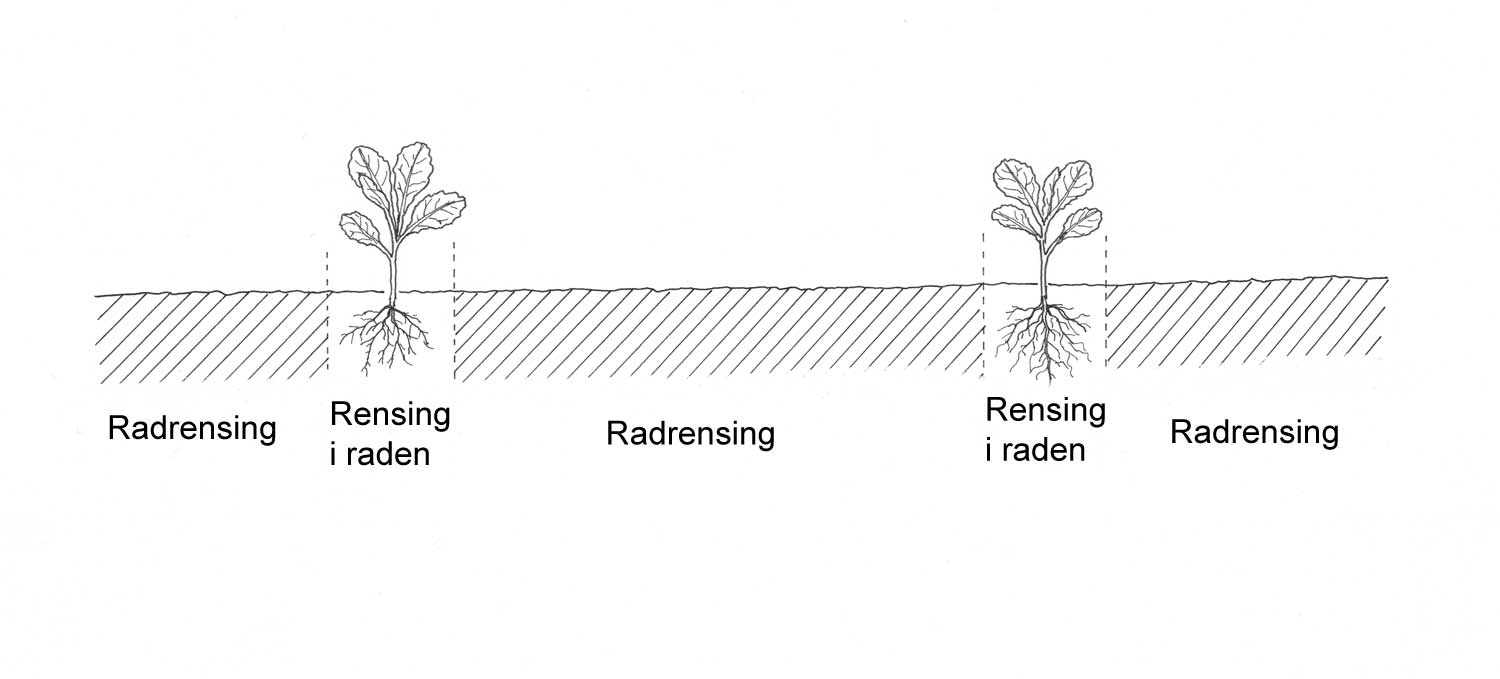
Arealet mellom radene blir å regne som helt åpne felt uten nytteplanter, og ugraskontrollen her blir nærmest å betrakte som brakking. Redskapsbredde og -utforming er da tilpasset slik at en ikke ødelegger kulturplantene i radene. Dette kalles fra gammelt av å radrense, i motsetning til å rense i radene (figur 4.12). Ugrasbekjempelse i radene, der i mot, vil vi her kalle å «rense i radene». Det er selvsagt mye vanskeligere å unngå at kulturveksten skades eller ødelegges når en renser i raden. Redskap for henholdsvis radrensing og rensing i raden kan kombineres på samme ramme slik at en kan gjøre begge deler på én kjøring.
I denne boka omtales de enkelte ikke-kjemiske bekjempningsmetodene, men siden de oftest ikke er så effektive som de kjemiske, er det viktig å være klar over at de ofte må kombineres med andre tiltak. Slike andre tiltak kan for eksempel omfatte forebyggende tiltak som jordarbeiding og utnytting av nytteplantenes konkurranseevne, vekstskifte og dekking mm, og som omtales andre steder i boka.
Manuell bekjempelse
Manuell bekjempelse inkluderer luking, dvs. at ugraset dras opp for hånd, hakking og skyfling. Hakka har et blad, vinkelrett på skaftet, som trykkes ned i jorda foran eller bak ugrasplantene og henholdsvis dras eller skyves mot plantene slik at de løsner (figur 4.13). Skyffelen (figur 4.13) løsner ugraset ved at et flattliggende jern skyves eller dras under ugrasplantene. På litt større planter kuttes eller trekkes røttene opp. Det vanligste er at luking og hakking/skyfling kombineres ved ugrasbekjempelse inne i radene, for eksempel ved at ugras tett inntil nytteplantene blir lukt, mens ugraset mellom plantene blir hakket eller skyflet. Luking vil også ofte være et nødvendig supplement når en bruker utstyr som renser i raden.
En form for manuell bekjempelse som i stor grad rasjonaliserer ugrasrenholdet, er hjulhakka. Den kan påmonteres blant annet skyffel og hyppeskjær (figur 4.13).
Mekanisk bekjempelse
Ved radrensing løsnes, rives opp, skjæres av eller overmoldes ugraset av faste eller roterende tinder, kniver og/eller ulike typer skjær eller børster som er montert på en ramme trukket av traktor eller hest eller skjøvet manuelt. De roterende redskapene monteres på horisontale, vertikale eller skråstilte aksler for henholdsvis vertikal, horisontal eller skrå rotasjonsretning. Driften av redskapene skjer med bakkedrift, dvs. at rotasjonen er en følge av jordmotstanden som oppstår når redskapet trekkes framover, eller fra traktoren via kraftuttaket. Redskaper som trekkes av hest eller skyves manuelt, har egen motor dersom arbeidsorganene skal drives. Ofte er dette elektriske motorer som drives fra et stort batteri. Redskap med bakkedrift kalles rulleredskap, og de med motordrift kalles rotorredskap.
Ikke-drevne rensere (figur 4.14): 
Tinde- eller skjærradrensere har gruntgående, faste tinder, skjær eller kniver som bearbeider hele jordarealet mellom radene. Vanlig ugrasharv brukes i noen tilfeller også i radkultur, men da arbeider den jorda både i og mellom radene.
Stjernehjulsrulle-radrensere eller harver har stjernehjul av metall på horisontale aksler for vertikal rotasjon med bakkedrift. De tar henholdsvis mellom radene og hele arealet.
Rad-fingerhjulrensere har bakkedrevne, roterende, tilnærmet horisontale hjul med gummifingre eller plastfingre på tilnærmet vertikale aksler. Fingrene går inn mellom kulturvekstene.
Rad-skrapepinnerensere, som også kalles torsjon- eller fjærpinnerensere, har runde, vinkelbøyde og vibrerende fastsittende stålspiler som løsner, skraper opp eller overmolder små ugrasplanter. Disse stilles som oftest for å ta ugras i radene.
Radrobotrensere er roboter med utstyr for å «se» forskjell på ugras og nytteplanter og spare de siste. Roboten luker ugraset eller dreper det mekanisk, kjemisk eller med stråling eller sterk varme/kulde.
Drevne rensere (figur 4.15):
Rotorradrensere har tinder på vertikale aksler for horisontal rotasjon (tilsvarer en seksjonsdelt rotorharv).
Seksjonsfreser har tinder eller kniver på horisontale aksler for vertikal rotasjon (tilsvarer en seksjonsdelt vanlig jordfres).
Børsterensere. Det finnes to typer, den ene har motordrevne, vertikale eller skråstilte aksler for horisontalt eller skrått roterende plastbørster, den andre har horisontalt liggende aksel med relativt lange plastbørster. Den siste bearbeider ofte jorda mellom radene også.
Ved ugrasharving drepes ugraset både ved nedgraving og oppriving. Best effekt vil man derfor få hvis det harves mens ugraset er smått, mest mulig på frøbladstadiet. Det er også viktig at det så langt som mulig, harves under tørre værforhold. Ugrasharving er en form for selektiv bekjempelse hvor selektiviteten består i at kulturplantene har større toleranse overfor overmolding og oppriving enn ugraset, bl.a. ved at de er større, mer opprette eller har dypere rotfeste. Ugrasharving virker derfor dårlig mot større frøugras og har liten eller ingen virkning mot rotugras når kulturplantene ikke skal skades for mye. Vanlig harvedybde er 2-3 cm. Virkningen på ugraset kan variere mye, men er ofte rapportert å ligge mellom 40 og 70 %, dårligst under fuktige forhold og best i tørt vær.
Ugrasharvene grupperes vanligvis på følgende måte:
- Seksjonsdelte stivtindharver har korte, stive tinder som den tidligere så kjente Korsmos ugrasharv. Denne harvtypen er bare aktuell før og like etter oppspiring av kulturveksten.
- Nettharver har bevegelige, mellomlange og stive enkelttinder. Disse er lenket sammen i et nett slik at enkelttindene er bevegelige både horisontalt og vertikalt når de møter motstand fra underlaget.
- Seksjonsdelte langtindharver (figur 4.16) har lange, runde og fjærende tinder, som enten er rette eller med knekk. Til en viss grad viker tindene til siden eller oppover/bakover når de møter økt motstand fra planteradene med korn av en viss størrelse. Slike harver er aktuelle i opptil 40 cm høye kulturvekster.
Termisk bekjempelse
Både sterk varme og sterk kulde skader cellemembranene i plantene slik at cellevæsken renner ut, og de overjordiske deler på plantene visner. Kuldebehandling har av ulike årsaker ikke blitt noen vanlig metode. Vi vil derfor konsentrere oss om varmebehandling, først og fremst i form av flamming, men også nevne litt om jorddamping. Andre termiske metoder omtales kort i (tekstboks 4.7).
Innen termisk ugraskontroll finnes flere ulike metoder både til flamming, damping og kuldenedvisning. Selv om de fleste av disse metodene ikke er i praktisk bruk i Norge i dag, vil vi her gi en kort oversikt.
Infrarød stråling
Den mest vanlige termiske metoden i dag er å drepe ugraset med en direkteflamme. En annen mulighet er å varme opp en overflate og bruke varmen som avgis fra denne til å drepe ugraset. I denne metoden varmer flammen opp keramiske overflater eller overflater av stål som sender infrarøde stråler mot plantene. Også deksler som brukes i forbindelse med vanlig flamming sender ut infrarøde stråler, men i så liten grad at slike flammere likevel regnes for å være direktevirkende. Det er ikke avklart om bruk av infrarød stråling er mindre energieffektivt enn direkte flamming. Ved infrarød stråling og i tette ugrasbestand «skygger» sannsynligvis ugraset mer for hverandre enn de gjør ved direkte flamming. Infrarød stråling er neppe brukbart til selektiv flamming i raden. Vi tror ikke at infrarød flamming får noen stor praktisk betydning, i alle fall ikke på kort sikt.
Gasstrøm oppvarmet av propanflamme
Tyske studier viser at uansett teknisk optimalisering, blir bare en del av den brukte energien nytta til selve flammingen. Resten går tapt til lufta. Konklusjonen i disse studiene var at videre reduksjon av energiforbruket bare kan skje ved å utarbeide et fullstendig nytt flammekonsept. Med dette som utgangspunkt ble prototypen «The Low Temperature Weeder» («LTW»), hvor en gasstrøm blir oppvarmet av en propanflamme, utviklet. Systemet er neppe egnet for selektiv flamming i raden.
Sprøyting med varmt vann opptil 100 oC kan brukes, men har lav kapasitet eller krever meget store deksler på utstyret slik at varmen holder seg lenge i plantene. I Australia brukes varmt vann for ugrasbekjempelse i parker o.l. på grunn av liten ulykkesrisiko, men det går med mye vann, og metoden blir for kostbar for vanlige jordbruksformål.
Sprøyting med varmt skum
I tyske undersøkelser er det gjort forsøk med bruk av varmt skum mot ugras i frukthager.
Kuldenedvisning
Ugrasbekjempelse ved å spre flytende nitrogen og «kullsyresnø», eller bare flytende nitrogen, utover ugraset har vært prøvd både i Sverige og Danmark. Metoden fungerte, men viste seg å være meget lite energieffektiv. Den var en kostbar metode og er bare aktuell i hager, parker, og andre steder der en må ta spesielle hensyn til brann- og eksplosjonsfaren ved flamming. Så langt vi vet, er det ikke noen som arbeider videre med denne metoden.


Metoder på eksperimentstadiet
Mange forskjellige tekniske metoder for ugraskontroll har vært foreslått i årenes løp. Noen av disse vil helt sikkert forbli på eksperimentstadiet, mens andre etter hvert kan bli vanlige. Bruk av høyspent strøm eller mikrobølger mot ugras er blant de nye metodene som for tida prøves ut (tekstboks 4.8).
Høyspent strøm
Høyspent strøm har vært prøvd i flere prosjekter. Til tross for lovende effekter på ugraset blir metoden av mange regnet for å være for farlig til å være aktuell.
Mikrobølger
Effekten på ugras av stråling med mikrobølger skyldes sannsynligvis først og fremst varmevirkningen. I motsetning til annen oppvarming trenger mikrobølgene inn i det mediet som skal varmes opp. En regner med at dette er av særlig betydning for å drepe ugrasfrø. Frøene må imidlertid være fuktige for at bølgene skal trenge inn. Dessverre må også den omliggende jorda varmes opp, slik at metoden er meget energikrevende. Dessuten kan det være risikabelt å rette bølgene ned mot jorda fordi de da kan reflekteres fra stein og bli kastet tilbake mot mennesker og dyr som går på jorda i nærheten. Den mest sannsynlige anvendelse vil være behandling av jord som blandes eller transporteres for bruk på plantefelt. Da kan behandlinga foregå inne i en trommel eller lignende.
Elektroporasjon
Elektroporasjon er basert på at cellemembranen i frø er ømfintlig for høyspente elektriske pulser eller støt. I undersøkelsene som er gjort, brukes pulser på 3000-6000 v/cm, og med varighet på brøkdeler av et sekund. Dette får elektrisk ladde molekyler i cellene til å bevege seg mot sin motpol slik at det oppstår permanente hull i cellemembranene og cellene dør. Innledende forsøk har gitt tilnærmet 100 % dreping av senneps- og rapsfrø plassert i henholdsvis vann og torvjord. Virkningen på ugrasfrø av smånesle, gjetertaske og meldestokk i naturlig jord var ca. 40 %. Det arbeides nå med å utvikle en feltmaskin for videre forsøk.
Skjærende vannstråle, laserstråling eller ultrafiolett stråling
Forskningsmiljøer som har studert virkningen av skjærende vannstråle konkluderer med at teknikken virker bra og har et stort potensiale. Ultrafiolett lys bedømmes som interessant, men det kreves betydelig forskningsinnsats før dette kan brukes i praksis. Laser vurderes som uinteressant på grunn av lav effektivitet og stor følsomhet for ulike forstyrrelser som oppstår ute på friland.
Vannstråleavskjæring er ingen ny teknikk i forskningen på ikke-kjemisk ugrasbekjempelse. Virkningen er trolig en momentan avklipping eller avriving. I undersøkelser i Sverige har et 0,25 mm munnstykke blitt rettet mot basis av plantene og en vannstråle med et trykk på 3000 bar har blitt sendt ut. Framdriftshastigheten på munnstykket langs planteraden har vært på opptil 18 km/time .
Ultrafiolett lys virker ved at de ytre delene av plantene oppvarmes sterkt. Skadebildet minner da også om det en ser ved flamming. Innledende forsøk har vist at det er forskjell på planteartenes ømfintlighet. Sannsynligvis passer denne bekjempningsmetoden best der det ikke er nytteplanter.
Robotbasert luking
Det arbeides med å utvikle robotbasert luking i flere land. I et europeisk prosjekt har en klart å få roboten til å skille smått ugras fra store kulturplanter og deretter fjerne en og en ugrasplante i raden ved hjelp av en roterende kniv. Hastigheten har vært 1 sekund per ugrasplante. Man ser for seg at metoden kombineres med vanlig radrensing mellom radene. Metodeutviklingen har kommet et stykke på vei, men er sannsynligvis ikke brukbart i praksis på en god stund.

Frontmontering og stabiliserende bakmontering gir ofte for unøyaktig styring til at det er mulig å radrense så nær planteradene som ved manuell styring. Resultatet blir fort at vi må bruke så mye mer arbeidstid (luking) eller gass til rensing i raden at det mer enn oppveier det en vinner på å spare operatøren på radrenseren. Midtmontering av redskap på traktoren gir best resultat, men er per i dag ikke veldig vanlige på norske bruk.
Noe det arbeides mye med nå, er videobasert, automatisk styreutstyr for bak- eller frontmontert radrenser. Et kamera som sitter på radrenseren tar løpende bilder, og en datamaskin tolker om raden hele tida er på riktig plass i bildet. Datamaskinen korrigerer radrenseren sideveis om nødvendig. Uansett system er det viktig at planteradene legges nøyaktig i forhold til kjøregangene og med nøyaktig avstand. Se for øvrig egen tekstboks 4.9 om ulike styringsutstyr.
Ulike miljøer har arbeidet med forskjellige systemer for styring av radrensere og annet utstyr. Generelt kan utstyret deles i to hovedtyper. Den første typen lager en får eller et spor i jorda som fungerer som utgangspunkt for styringa. Den andre typen er videobasert.
Får eller spor i åkeren
I Sverige har slikt mekanisk styringsutstyr (JT) allerede vært i salg en tid. Det lages en får i bakken ved såing eller planting. Ved radrensing senere går et følerhjul i fåra og gir signaler til et elektrohydraulisk system på radrenseren når denne skal forandre kurs i forhold til fåra.
I Norge har det blitt utviklet et liknende utstyr. En følersko i fåra styrer redskapet ved hjelp av et enkelt elektrohydraulisk system. Følersko ble valgt istedenfor hjul i dette tilfellet, fordi det ofte er mye stein i norsk jord. Om utstyret blir tilgjengelig for salg, gjenstår å se.
En ulempe ved systemer med styrefår er at fåra lett blir ødelagt av tungt regn, av mekanisk radrenserutstyr og lignende.
Et annet mekanisk, norsk system (utviklet av Harald Lægereid, Lærdal/Arendal) er basert på at dype kjøreganger blir brukt som styrespor for traktor og eventuelle bærehjul på det bakmonterte utstyret. Det følger også med ekstrautstyr for å sikre at hjulsporene som kjøres opp før såing eller planting, blir lagt riktig. Utstyrer har fungert bra, men på stiv jord kan det være et problem at vann blir stående i de hardkjørte hjulsporene i regnværsperioder.
Videobaserte systemer
Flere typer utstyr er allerede på markedet, for eksempel fra to danske produsenter, ECO-DAN og «Frank Poulsens Autopilot», og det britiske Garford.
Vi skal her gå litt nærmere inn på utstyret fra ECO-DAN. Dette styresystemet er utviklet for å kunne monteres på både nye og eksisterende redskaper, og kan styres etter både planteraden og jordspor. På radrenseren monteres utstyr som gjør at den kan forskyves sideveis ved hjelp av en hydraulisk sylinder. Et såkalt «vision kamera» er ryggraden i styresystemet. Det tar bilder fortløpende av planteraden som skal følges. Selve «hjernen» i systemet er en datamaskin med tilhørende programvare som sørger for styring av radrenseren ut fra bildene som blir tatt. Når datamaskinen tolker at radrenseren ikke er riktig plassert i forhold til raden, sender den strøm til en elektisk styrt ventil som igjen sender olje til den hydrauliske sylinderen som skyver radrenseren til side. Systemet har en nøyaktighet på pluss-minus 2 cm selv om det kjøres 8 km/t.
I en del vekster er det viktig å starte ugrasbekjempningen tidlig. Derfor er det utviklet et spesialkamera med innebygd laser, som i tillegg til å følge planteraden, også kan følge et styrespor. Styresporet lages samtidig med såing, og radrenseren kan følge dette i tida før kulturveksten er synlig som rad.
Med selektiv flamming i raden i voksende kulturplanter utnytter man forskjeller mellom kulturvekstens og ugrasets høyde og varmetoleranse. Selektiv flamming vil altså si at man flammer i en situasjon hvor kulturplanter og ugras står side om side, og at behandlinga i størst mulig grad bare skader og dreper ugrasplantene. Samtidig er det viktig at en mest mulig unngår å treffe kulturplantens vekstpunkt. Selektiv flamming gjøres mest skånsomt med to skråstilte brennere (se figur 4.20), montert på en vogn som trilles eller på ei ramme bak en traktor. Stikkløk og potet kan også flammes selektivt under deksel. Ved Institutt for matematiske realfag og teknologi, UMB, har metoden tidligere blitt videreutviklet ved hjelp av to åpne brennere («ITF-brenneren»). Denne gir en bred, kort og forholdsvis tynn flamme som er ideell for slik flamming, og brenneren skal stilles med munningen ca. 5 cm over bakken. Denne metoden er undersøkt i mange planta grønnsakvekster. Alt etter grønnsakslag og værforhold tålte disse vekstene doser på opptil 5-9 kg gass per dekar. Virkningen på ugraset økte imidlertid for doser opptil 11 kg/dekar. Selv om man altså har relativt gode erfaringer med slik flamming, bør man prøve seg litt fram på hvert enkelt sted før en flammer store arealer selektivt. Har en utstyr som trilles manuelt, kan en endre dosen ved å gå raskere eller saktere. Det samme gjelder jo også traktormontert utstyr der dosen endres vha kjørehastighet.
Forsøk med selektiv flamming under deksel har blitt utført i stikkløk. På grunn av sen ugrasspiring ble det ikke flammet før løkspirene var 5-10 cm høye. Dette gav noe nedsatt vekst på løken for doser over 3 kg/dekar. I andre tilsvarende forsøk, hvor det ble brukt vanlig metode med to åpne brennere, fikk en ikke skader før ved doser over 9 kg/dekar. Også i den flerårige urten peppermynte har det blitt prøvd å flamme både med to åpne, skråstilte brennere (som gav godt resultat med dose 5 kg/dekar) og under deksel (som gav store skader ved 5 kg/dekar). Ut fra disse resultatene må selektiv flamming under deksel frarådes dersom den ikke er prøvd og funnet akseptabel, på den aktuelle vekst og det aktuelle stedet på forhånd.
Hvordan flammeutstyret er oppbygd er avgjørende for virkningen på ugraset. Den teknisk interesserte vil finne mer omkring brennerplassering, brennertype og dekseltyper i tekstboks 4.10.
Brennerplassering og brennertyper (figur 4.20, A, B og C)
Undersøkelser har vist, og det gjelder uansett om det benyttes deksel eller ikke, at brennerne bør plasseres fremst og flamme bakover. Når det gjelder valget mellom runde brennere eller flate, brede brennere, vil det være avhengig av hvilken brennervinkel (vinkel mellom brenner og jordoverflaten) som benyttes. Ved flatstilte brennere (liten vinkel mellom brenner og jordoverflate) vil runde brennere gi bedre virkningsgrad enn flate, brede brennere. Ved mer brattstilte brenner (dvs. større brennervinkel) varierer ikke virkningsgraden mellom disse brennertypene i noen særlig grad.
At brennervinkelen påvirker effekten av brennertypene, skyldes at når brenneren er flatstilt, vil runde brennere få større rekkevidde langs kjøreretningen og dermed også lenger virkningstid per ugrasplante. Ved mer brattstilling av brennerne blir forskjellen i rekkevidde mellom runde og flate, brede brennere mindre. Ved selektiv brenning vil de brede, flate brennerne rekke over flere planter på tvers av kjøreretningen enn de runde. Dette er viktig ved selektiv flamming (se figur 4.20, A og B).
Gasstilførsel, gasstrykk og avstand mellom brenner og ugras
I laboratorieundersøkelser har man sett på ulik gasstilførsel, gasstrykk og avstander mellom brenner og ugras.
Økt gasstilførsel, fra 1,2 til 3,4 kg per time førte til:
Økende avstand mellom brenner og nedslag på bakken førte til:
Økende gasstrykk ved samme gasstilførsel førte til:
Bruk av deksler og andre tiltak for å øke virkningen på ugraset
Hensikten med å bruke deksler ved flamming er todelt: (a) Beskytte mot uønsket direktekontakt mellom nytteplanter og flamme, for eksempel ved termisk radrensing, men også sikkerhetsmessig mellom flamme og operatør. (b) Øke virkningen mot ugraset, grunnet bedre kontakt mellom flamme og ugras, dessuten lenger behandlingstid. Både gass, varmluft og avgasser søker opp fra plantene hvis det ikke er noe stengsel.
Noen eksempler på tiltak for å øke bekjempingseffekten:
Litt om økonomi, brennerkapasitet, dose og kjørehastighet
Arbeidskostnader og investeringskostnader er oftest langt større enn gasskostnadene. Spesielt på større bruk innebærer dette at kjørehastighet og arbeidsbredde er viktige faktorer.
I og med at en må opp i et visst energiforbruk for å oppnå et gitt flammingsresultat, er det viktig å forstå energiformelen ∑ Energi = (Temperatur x tid x konstant), dvs. at energi er lik summen av de ulike temperaturer en ugrasplante blir utsatt for, multiplisert med tida som hver av disse temperaturene virker i, igjen multiplisert med en konstant. For å oppnå en viss energiinnsats ved flamming kreves altså høyere temperatur ved stor enn ved liten kjørehastighet. Høyere temperatur oppnås ved å øke tilført gassmengde forutsatt blant annet tilstrekkelig økning i brennerkapasitet og lufttilførsel. Har lufttilførsel og rom under dekslene vært rikelige i forhold til brennerkapasiteten, kan større kjørehastighet oppnås ved å senke dekselhøyden eller øke deksellengden. Her kommer det også inn at økt kjørehastighet gir økt luftstrømning under dekslet, slik at økt gasstilførsel tåles bedre.
Ved selektiv flamming brukes som regel ikke deksel. Med brede, flate brennere med liten rekkevidde framover, oppnås likevel flamming av lange deler av raden når brennerne monteres i «selektivstilling» dvs. med en brenner montert på hver side av raden og tver på denne (se figur 4.20 A og B). Ved å flamme flere rader samtidig øker kapasiteten. Dette betyr imidlertid oftest høyere investeringskostnader enn det en øking i kjørehastigheten representerer.
Termisk kontroll
Regulering av temperatur kan brukes på mange måter. Kanskje er temperaturens påvirkning på mikroorganismer noe av det vi bruker hyppigst i dagliglivet uten å tenke så mye over det. Hver dag putter vi ting inn i kjøleskapet, ikke bare fordi det er godt med kald melk, men fordi vi ønsker å forlenge holdbarheten, dvs. hemme vekst av mikroorganismer. Melken er på forhånd pasteurisert, dvs. oppvarmet til 70 °C for å drepe skadelige bakterier. I noen tilfeller er det til og med slik at de patogene eller uønskede mikroorganismer er mer sensitive for varme enn en del andre mer ønskede mikroorganismer, men mikrobenes optimale, maksimale og minimale temperaturområder varierer selvsagt svært mye. Lave temperaturer og kulde vil ofte hemme veksten til mange sopp og bakterier, men uten å drepe dem. Faktisk kan mange overleve fint i nedfrosset tilstand over ganske lang tid. Mange mikroorganismer har egne overlevelsesorganer for å klare lave temperaturer eller tøffe perioder i sin livssyklus. Høye temperaturer er det derimot verre å takle for de uønskede små organismene, og det finnes mange eksempler på bruk av varme for å kvitte seg med spesielle patogene sopp eller bakterier. Oppvarming av jord, og varmebehandling av småplanter, stiklinger, settepoteter eller frø er utprøvde metoder for dette, men med litt ulike strategier og litt ulik grad av selektivitet.
Oppvarming av jorda ved hjelp av sol, damp eller varmt vann har vært prøvd og brukt i mange sammenhenger både i regulert klima (veksthus) og på friland. Metodene er imidlertid energikrevende, og er ikke selektiv mhp. Andre mikroorganismer i jord. Likevel kan metodene ha sine fordeler, kanskje særlig i konvensjonelt jordbruk dersom man har vanskelige patogene, jordboende mikroorganismer hvor det eneste alternativet er svært giftige kjemikalier (jorddesinfeksjonsmidler brukes i enkelte andre land, men er ikke tillatt i Norge). Varmebehandling av frø ved hjelp av varmt vann eller damp, er kanskje den mest aktuelle termiske kontrollmetoden innen økologisk jordbruk.
Fysisk kontroll av sjukdommer
Betegnelsen fysisk kontroll av sjukdommer kan fort overlappe med enkelte forebyggende tiltak mot sjukdommer. Tiltak under denne betegnelsen vil derfor bare nevnes kort her. Fysisk kontroll av sjukdom kan for eksempel være å hindre spredning av smittestoff gjennom å rydde og fjerne syke planter eller planterester som sjukdom kan overleve på. Fjerning av infiserte blader under epletrær for å hindre spredning av epleskurv er eksempel på dette. Fjerning av halm for å begrense smitte fra halmrester til kornplanter vil dermed også kunne kalles fysisk kontroll av sjukdom. I noen tilfeller er det aktuelt å fjerne planter som er vertsplanter for særlig destruktive sjukdommer. Dette er høyst aktuelt når det gjelder den alvorlige sjukdommen pærebrann og dens alternative vertsplanter kulemispel og bulkmispel. Knusing av potetris kan forebygge smitte av potet-tørråte fra riset og ned i knollen, og kan kalles fysisk tiltak.

Fysisk kontroll
Når vi snakker om fysisk kontroll av skadedyr tenker de fleste med en gang på fluesmekker! Dette er en form for fysisk kontroll som er meget effektiv, men kanskje ikke så veldig praktisk i stor skala. Prinsippet er likevel det samme som når vi pløyer eller harver og knuser skadedyrene som befinner seg i jorda.
Andre former for fysisk kontroll er ulike metoder for utestenging av skadedyret. Det som er best kjent og mest utbredt er bruk av fiberduk eller insektsnett som legges over kulturen slik at flyvende skadedyr på søken etter vertsplanter ikke kommer til (figur 4.21).
Det er en forutsetning at skadedyret ikke er på plass når kulturen dekkes over, f.eks. fra overvintring (uten vekstskifte) eller tidlig angrep. Hvis kulturen er dekket over slik at det ikke finnes inngangsmuligheter for skadedyret, kan denne metoden være svært effektiv. For noen kulturer vil klimaendringen under duk/nett være gunstig for vekst, for andre ikke, f. eks. kålrot (figur 4.22).
En ny metode som også baserer seg på utestenging av skadedyret, men uten merkbar klimaendring, er gjerder av insektsnett. Ved første øyekast virker det ulogisk å stenge ut flyvende skadedyr med et gjerde, men metoden bygger på at flere skadedyr, f.eks. kålflue, flyr lavt over bakken i søken etter vertsplanter. Dette gjør at de støter på gjerdet på vei inn i åkeren. Gjerder som er prøvd mot kålfluer er satt opp i 150-180 cm høyde. På toppen er nettingen brettet over og mot utsiden av åkeren (figur 4.23). Når en kålflue lander på en loddrett flate går den oppover mot sollyset. Når den har landet på gjerdet, går den oppover og støter på toppen som er brettet slik at flua ikke kommer videre. Da snur den og går fra side til side. Det er unaturlig for flua å gå nedover. Derfor blir den fanget og vil etter hvert bli utslitt, utsultet eller fly ut fra gjerdet og enten fly ut av området eller snu og støte på gjerdet igjen og gjenta hele prossessen. Denne metoden har i flere forsøk vist seg å være svært effektiv. Når det har vært mye angrep innenfor et gjerde har det som regel vært i nærheten av høy vegetasjon, i kupert terreng eller lave punkter eller andre svakheter i oppsetting av gjerdet (figur 4.23). Som nevnt er metoden mest utprøvd mot kålfluer, men andre skadedyr som flyr lavt vil også være hemmet av et gjerde, f.eks. gulrotflue, jordlopper og noen tege- og snutebillearter.
Termisk kontroll
Varmebehandling har lenge vært brukt til desinfisering av planter til utplanting, f.eks. jordbærplanter som er infisert med midd. Både egg og voksne midd blir drept av oppvarming til mellom 42 og 50 °C i noen minutter (tabell 4.5).
Tabell 4.5 Temperatur og nødvendig behandlingstid for
å drepe jordbærmidd i småplanter av jordbær (Stenseth 1975).
| Temperatur °C | Tid (minutter) |
| 42 | 24,0 |
| 44 | 12,0 |
| 46 | 8,5 |
| 48 | 6,0 |
| 50 | 4,8 |
Også nematoder blir drept ved temperaturer omkring 46 °C. For jordbærplanter kan dette være aktuelt for stengelnematode og bladnematode som drepes etter hhv. 7 og 10 minutter ved denne temperaturen.
Interessen for alternative plantevernmidler er økende hos mange økologiske bønder. Årsaken til interessen kan enten være at man i dag ikke har andre tilstrekkelig gode tiltak, og at produksjonen derfor er risikabel, eller at dagens metoder er for arbeidskrevende og upraktiske. Et eksempel på det siste kan være bruken av fiberduk eller insektsnett for skadedyrkontroll i grønnsaker. Dette er tidkrevende både ved førstegangs utlegging og ved ugrasrenhold senere i vekstsesongen. I det konvensjonelle landbruket er det også økende interesse for alternative plantevernmidler, dels fordi mange av de gamle midlene ikke lenger er tillatt, og dels fordi mange mener at alternative plantevernmidler er mer miljøvennlige enn de tradisjonelle.
Det er gjort litt med «alternative plantevernmidler» eller «plantevernmidler av naturlig opphav» også i Norge, men lite i forhold til hva som er gjort i mange andre land. Både fordi vi i Norge ikke ønsker at økologisk landbruk bare skal være en «grønn» utgave av konvensjonelt landbruk, men også fordi vi i dag ikke vet nok om virkningen av slike på skadegjørere og miljøet, skal vi ikke bruke veldig mye plass på alternative plantevernmidler her.
For kontroll både av sjukdommer og skadedyr kan stoff som dreper skadegjøreren, eller i hvert fall reduserer utviklingen av skadegjøreren, være nyttig. Stoff som styrker kulturplantenes forsvarsmekanismer kan også være interessant for begge gruppene av skadegjørere. For skadedyrkontroll er i tillegg stoff som forstyrrer skadedyrets søken etter vertsplante eller make, en fascinerende og aktuell tilnærmelse.
Listene over preparater som er tillatt brukt i økologisk planteproduksjon, og likeledes planter o.a. som er tillatt brukt til framstilling av hjemmelagde preparater, kommer vi tilbake til i kulturheftene.
Det har generelt vært en restriktiv holdning i Norge angående bruk av «grønne» plantevernmidler. Hvis slike midler skal godkjennes i Norge, må de ha en avgjørende betydning for kontrollen med visse skadegjørere, slik at ikke hele produksjonen går tapt. Siden vi tross alt har flere tiltak tilgjengelig mot ugras, er ikke behovet for alternative midler særlig påtrengende.
Direkte bekjempelse i form av kjemiske plantevernmidler har vært og er noe man forbinder med konvensjonell dyrking. I en god del tilfeller er det likevel behov for direkte bekjempelse av plantesjukdommer, selv om alle forebyggende tiltak er forsøkt brukt. Kobber er eksempel på et middel med svært lang historie innen bekjempelse av plantesjukdommer, som også er brukt ved økologisk dyrking. Kobbersulfat ble introdusert som beisemiddel i korn allerede i 1761. Det finnes en rekke planteekstrakter og andre naturlig framstilte midler med mer eller mindre beviselig effekt mot ulike sjukdommer, men disse midlene er ikke alltid godt nok undersøkt. Et annet problem er at midlene er vanskelige å få godkjent, ettersom kravene til dokumentasjon er de samme enten middelet har opprinnelse i et naturlig forekommende stoff eller er framstilt kjemisk. Ofte finnes det ingen store firmaer som har økonomisk interesse av å utvikle slike midler til bruk i økologisk landbruk. Det foregår imidlertid forskning og undersøkelser på effekten av alternative midler. Midlene vil nok i første rekke rette seg mot de sjukdommene som vanskelig lar seg bekjempe godt nok med forebyggende tiltak. Tørråte i potet og gråskimmel i jordbær vil kunne være områder der man trenger alternative midler for å holde sjukdommen i sjakk.
Det finnes få direkte bekjempelsesmidler som er godkjent til bruk mot sjukdommer i økologisk dyrking. De meste benyttede midlene generelt i Europa baseres enten på kobber eller svovel. Svovelpreparatet «Thiovit» er godkjent og brukt mot skurv i økologisk fruktdyrking i Norge. Svovelkalk var mye brukt tidligere, men er ikke lenger tillatt brukt i Norge. Svovelkalk er fortsatt mye brukt i andre land i Europa, og er i flere land det viktigste preparatet mot sjukdommer i økologisk fruktdyrking. Preparater som inneholder kobber finnes i flere former. Kobberpreparater er også mye brukt i fruktdyrking i andre europeiske land. Det er imidlertid et klart mål innen EU å redusere bruken av kobber generelt. Det finnes ulike kobberpreparater, deriblant kobberkalk (kobberoksyklorid) som også er brukt i Norge. Kobber har god forebyggende effekt mot skurvsjukdommer i kjernefrukt og er også benyttet mot bakteriekreft i steinfrukt. Kobber har dessuten brukbar effekt mot tørråte i potet. Det finnes reguleringer i flere land for hvor store mengder kobber som er tillatt brukt per arealenhet og år. Kobberpreparater er ikke tillatt brukt i norsk økologisk dyrking. Ulempen med kobber er at det ikke brytes ned, men akkumuleres i jord, avhengig av utgangsnivået i jorda, mengda i gjødsel, jordas pH-nivå og bufferkapasitet, avrenning og uttynning ved hjelp av nedbør og vanning, samt kobberinnhold i middelet som brukes.
Per nov. 2005 finnes tre biologiske midler som er godkjent mot sopp: Cedomon, Mycostop og Rotstop. Alle tre midlene er basert på levende organismer og prinsippene er beskrevet i underkapitlet Biologisk kontroll av plantesjukdommer. Cedomon (basert på bakterien Pseudomonas chlororaphis) er godkjent som beisemiddel mot frøoverførte sjukdommer i bygg og havre, mens en variant av middelet (Ceral) er utviklet til bruk i hvete. Mycostop er basert på jordbakterien Streptomyces griseoviridis. Den er virksom mot både frøoverførte sjukdommer og andre sjukdommer. Den er godkjent i en del veksthuskulturer, og er først og fremst anbefalt mot visse arter av Pythium, Fusarium, Alternaria samt Phomopsis. Rotstop er et middel bestående av sporer fra soppen Phlebiopsis gigantea . Middelet brukes på trestubber for å hindre infeksjon og spredning av rotråtesopper.
Utenom de godkjente preparatene mot ulike sjukdommer, finnes en rekke alternative midler som tilskrives en viss sjukdomshemmende effekt. En del av disse midlene er kun på forsøksstadiet, eller de er ikke prøvd i forsøk i det hele tatt. En del inneholder komponenter som i forsøk har vist seg å ha en viss effekt mot sjukdommer. Flere midler som er omtalt som plantestyrkende midler eller komposteringsmidler, kan nok påvirke andre mikroorganismer, deriblant sjukdomsorganismer. Disse er imidlertid ikke godkjent til bruk mot sjukdommer, og er som regel ikke testet i forsøk. Effekt mot skadegjørere kan selvfølgelig ikke utelukkes. Komposteringsmidler, som for eksempel EM = effektive mikroorganismer og Terra Biosa, kan også påvirke mikrofloraen, og dermed også påvirke sjukdomsorganismer. Det vil være aktuelt å prøve ut flere slike preparater i framtida.
Ulike midler basert på alger er testet for sjukdomshemmende effekter i flere land. Dette er preparater som inneholder flere ulike komponenter, og innholdet (og dermed også effektene) kan variere mye mellom preparatene. Også jordforbedringsmidler, som rekemel, har komponenter (kitosan) med potensiell sjukdomshemmende effekt. Resultater fra forsøk varierer fra ingen effekt til bra effekt, avhengig av en rekke forhold, bl.a. typen middel og skadegjører.
Kompost og preparater fra kompost, som for eksempel «kompost-te», er også vist å ha sjukdomshemmende effekter i mange forsøk innenlands og utenlands. Kompost kan påvirke sjukdomsorganismer på en rekke måter; for eksempel pga endring av jordstruktur, næringstilgang eller vannhusholdning. Spesielt to biologiske mekanismer bak sjukdomshemmende effekter er blitt viet mye oppmerksomhet: antagonisme og indusert resistens. Kompost kan inneholde mikroorganismer med antagonistiske effekter mot sjukdomsorganismer (se for øvrig avsnitt om «Dødt jorddekke», samt avsnitt organisk gjødsel og sjukdommer). Indusert resistens oppnås når plantenes forsvarsmekanismer blir stimulert og kan motstå et sjukdomsangrep (med mindre skade som resultat). Ikke alle typer kompost har sjukdomshemmende effekt. Visse komposter kan til og med favorisere sjukdomsutvikling. Beising mot frøoverførte sjukdommer er et område innen økologisk dyrking der behovet for direkte bekjempelsesmidler er tydelig til stede og alternativene få. Alternative beisemidler i form av for eksempel planteekstrakter og andre midler (vurdert som uskadelige for miljø og mennesker) blir derfor undersøkt og vurdert i mange land.
Som vi allerede har vært innom, er det svært mange forhold som påvirker skadedyrenes atferd. En side av dette er et fagområde som ofte kalles «kjemisk økologi». Her er det snakk om naturlig forekommende stoffer, både stoff som insektene produserer selv, og stoffer som vertsplanter produserer, og hvordan slike stoff påvirker skadedyrenes atferd. Dette «kjemiske språket» mellom insekter og mellom insekter og planter, er svært komplisert, og mange muligheter for utnyttelse til skadedyrkontroll vil helt sikkert bli prøvd ut i årene framover. Hittil har spesielt to muligheter fått stor oppmerksomhet:
Per i dag har vi ingen slike stoff tilgjengelige for skadedyrkontroll ved økologisk dyrking i Norge. Et lite unntak er luktstoffet som brukes for varsling av skadedyrangrep, brukt i kålfluefeller. I utgangspunktet er disse insektfellene brukt for å varsle bonden om når han skal sprøyte, men kan også brukes for å bestemme når fiberduk/insektnett skal legges på en kultur.
I andre europeiske land er ulike stoffer godkjente og benyttes ved økologisk produksjon. Et eksempel er bruk av forvirringsteknikk med feromondispensere for å forstyrre parring hos eplevikler og plommevikler. I Sveits er dette i dag en standard metode i større hager, og det er vanlig med en dispenser i annethvert tre. Denne forvirringsteknikken fungerer best ved store sammenhengende arealer, og når det er åpne jorder eller enger rundt frukthagen.
Selv om vi i Norge ikke i dag har slike stoffer eller teknikker tilgjengelige, arbeides det med dette emnet i norske forskningsprosjekter. I ett prosjekt undersøkes luktstoff som kan forstyrre jordbærsnutebillen ved enten å virke avskrekkende («repellenter») eller tiltrekkende («attraktanter») (tekstboks 4.11).
For å utvikle nye plantevernmetoder mot insektskader innen økologisk landbruk forutsettes det at man har inngående kunnskap om de insekt-planterelasjoner som man ønsker å påvirke. Oftest er disse relasjonene kompliserte systemer med mange variabler både med tanke på insektenes evne til å lokalisere vertsplanter, og plantenes evne til å motstå insektskader. Uten kunnskap om disse forholdene vil utfordringen med å utvikle hensiktsmessige og effektive plantevernmetoder bli uoverkommelig. Enkelttiltak viser seg ofte å være utilstrekkelige, så derfor bør en forske på flere fronter for å kunne kombinere ulike metoder for å få best mulig beskyttelse. En innfallsvinkel er å påvirke insektenes atferd ved enten å lokke dem til feller eller å få dem til å unnvike nytteplantene. I den forbindelse vil bruken av duftstoffer fra vertsplantene være sentral.
Et pågående prosjekt ved Bioforsk har med bakgrunn i nevnte problemstilling til hensikt å utvikle en plantevernmetode mot skader fra jordbærsnutebillen basert på substanser jordbærplantene produserer. Skadene som plantene påføres av jordbærsnutebillen skjer når hunnbillen snitter stilken til blomsterknoppene etter egglegging. Knoppene tørker inn og vil etter en tid falle av.
For å nå prosjektets målsetninger er det blitt foretatt innsamling og omfattende kjemiske analyser av duftstoffer fra jordbærplanter. Resultater fra disse forsøkene viser bl.a. at i jordbærplantene finnes en rekke ulike alkoholer, aldehyder, estere, terpener og andre aromatiske forbindelser. For å identifisere hvilke av disse duftene jordbærsnutebillen oppfatter er det blitt utført elektrofysiologiske registreringer fra antennene til insektet. Ved slike registreringer er det mulig å finne ut hvilke duftstoffer som utløser en fysiologisk respons i lukteorganet til billen. På denne måten er det blitt identifisert en rekke komponenter fra jordbærplantene som trolig har betydning for billens vertsvalg. Men om man identifiserer hvilke stoffer et insekt sanser sier det ingen ting om hvilken effekt stoffene har på billen: Om de er tiltrekkende eller frastøtende. Derfor er det iverksatt omfattende atferdsforsøk. Imidlertid må studiene suppleres med både flere laboratorieforsøk og ikke minst flere feltforsøk før man kan trekke noen konklusjoner. Om man finner enkeltstoffer eller en blanding av flere duftstoffer som påvirker billens atferd i felt, så gjenstår det å utvikle en funksjonell metode for anvendelse av resultatene i praksis. Hvordan substansene skal brukes avhenger av hvordan de påvirker billens atferd.
Rognebærmøll er en liten sommerfugl som har rogn som vertsplante. Møllen legger egg på rognebær, og larvene utvikler seg inne i bærene. Rogn har imidlertid utpreget vekselbæring. Det vil si at bærmengda på rogn varierer mye fra år til år, og det er bare i år med lite bær på rogna at møllen angriper eple. Møllen er altså i stand til å velge mellom rogn og eple.
Fordi rognebærmøll er nattaktiv og må finne fram i blandet skog, er det sannsynlig at luktesansen er viktigere enn synet når møllen skal velge sted for å legge egg. Møllen må altså kunne lukte forskjell på rogn, eple og andre treslag.
Det vi kaller lukt er en blanding av flyktige kjemikalier i luften. Ved Bioforsk har vi i samarbeid med kjemikere ved SLU i Sverige, samlet lukt fra frukter av rogn og eple, og vist at møllen reagerer på flere av disse luktkomponentene. Vi har også kunnet vise at møllens reaksjon på noen av stoffene er positiv, det vil si at møllen tiltrekkes av lukten.
Det har lenge vært kjent at insekter kan lokkes til feller ved hjelp av lukt. Det er imidlertid som oftest hunnenes kjønnsferomon som er brukt, og slike feromonfeller fanger bare hanner. Ved å bruke rognlukt burde det derimot være mulig å fange rognebærmøllhunnene før de legger egg, eller «lure» dem til å legge eggene et annet sted enn på eple, selv om de ikke finner rognebær. Innledende feltforsøk har vist at en blanding av to av de luktstoffene vi har identifisert tiltrekker både hunner og hanner i betydelig grad. Antagelig er det mer komplisert enn som så, og en «super rognlukt» er trolig en blanding av mange stoffer i et bestemt mengdeforhold. Det kan også bli en utfordring å finne en god formulering, det vil si at stoffene må fordampe med jevn hastighet over tilstrekkelig lang tid.
Kromatogrammet (figur 4.25) viser noen av luktstoffene fra rogn og eple. Rognebærmøll har i forsøk reagert på de åtte stoffene som er markert med røde stiplete linjer. Legg merke til at det er tydelig forskjell på den mengdevise sammensetning av disse komponentene i rogn og eple. Rogn og eple lukter forskjellig!

Ascard, J. 2003: Ogräs och ogräsreglering i ekologisk grönsaksodling. Jordbruksverket, Jordbruksinformation 21, 23s.
Baker, K.F. 1980: Microbial antagonism – the potential for biological control. I: Contemporary Microbial Ecology, D.C. Ellwood, J.N. Hedger, J.M. Latham, M.J. Lynch & J.H. Slater (red.). Academic Press, London, s. 327-347
Barbosa, P. 1998: Conservation biological control. Academic Press, San Diego, 396 s.
Bedding, R.A., R. Akhurst & H. Kaya 1993: Nematodes and the biological control of insect pests. Australia, CSIRO, s. 178
Bellows, T.S. & T.W. Fisher 1999: Handbook of biological control. Academic Press, San Diego, 1046 s.
Bengtsson, M., G. Jaastad, G. Knudsen, S. Kobro, A-C. Bäckman, E. Petterson & P. Witzgall 2005: Plant volatiles mediate host switch in apple fruit moth, Argyresthia conjugella. Entomologia Experimentalis et Applicata (under trykking).
Boland, G.J. & L.D. Kuykenhall (red.) 1998: Plant-microbe interactions and biological control. Marcel Dekker, Inc, New York, 442 s.
Butt, T.M., C.W. Jackson & N. Magan (red.) 2001: Fungi as biocontrol agents, progress, problems and potential. CABI Publishing, Wallingford, UK, 390 s.
Edvardsen, O.E. 2002: Gjerde mot kålflua - framtidas løsning? Grønn forskning 2, s. 123-1127
Gaugler, R. (Ed) 2002: Entomopathogenic nematology. UK, Cabi publishing, 388 s.
Gaugler, R. & H.K. Kaya, (red.) 1990: Entomopathogenic nematodes in biological control. Boca Raton, FL, CRC Press, 356 s.
Gurr, G. & S. Wratten 2000: Biological control: Measures of success. Kluwer Academic Publishers, Dordrecht, 429 s.
Hajek, A.E. & R.J. St. Leger 1994: Interactions between fungal pathogens and insect hosts. Annual Review of Entomology 39, s. 293-322
Haukeland, S. 1993: Entomopathogenic nematodes found in Norway. Norwegian Journal of Agricultural Sciences 7, s. 17-27
Johansen, T.J. & L.S. Toften. 2003: Nettgjerder begrenser angrep av kålfluer. Norden 2003, s. 30-31
Klingen, I., S.H. Salinas & R. Meadow 2002: Checklist of naturally occurring pathogens of insects and mites in Norway. Norwegian Journal of Entomology 49, s. 23-28
Malais, M.H. & W.J. Ravensberg 2003: Knowing and recognizing. The biology of glasshouse pests and their natural enemies. Koppert B.V. R&P.C. Quimby 2001: Evaluating indirect ecological effects of biological control. CAB Publishing, Wallingford, 261 s.
Plantevernleksikonet er en nettbasert tjeneste som omfatter informasjon om biologi og bekjempelse av skadegjørere, samt informasjon om en del nyttedyr. Plantevernleksikonet er gratis og uten forpliktelser for brukeren. Tjenesten er utviklet av NIBIO Divisjon bioteknologi og plantehelse. Plantevernguiden er en integrert del av tjenesten. Drift, oppdatering og videreutvikling av Plantevernleksikonet finansieres av handlingsplanmidler fra Landbruksdirektoratet og kunnskapsutviklingsmidler fra Landbruks- og matdepartementet. Bilder i Plantevernleksikonet kan kopieres og brukes dersom de er fra NIBIO-/Bioforsk-/Planteforsk-ansatte, og det refereres til rett kildehenvisning, f.eks.: "Foto: ... fra Plantevernleksikonet, E. Fløistad, NIBIO".
NIBIO har ikke økonomisk ansvar for tap som måtte oppstå ved bruk av tjenesten.
Plantevernleksikonet © 2026 NIBIO
Plantevernleksikonet er en nettbasert tjeneste som omfatter informasjon om biologi og bekjempelse av skadegjørere, samt informasjon om en del nyttedyr. Plantevernleksikonet er gratis og uten forpliktelser for brukeren. Tjenesten er utviklet av NIBIO Divisjon bioteknologi og plantehelse. Plantevernguiden er en integrert del av tjenesten. Drift, oppdatering og videreutvikling av Plantevernleksikonet finansieres av handlingsplanmidler fra Landbruksdirektoratet og kunnskapsutviklingsmidler fra Landbruks- og matdepartementet. Bilder i Plantevernleksikonet kan kopieres og brukes dersom de er fra NIBIO-/Bioforsk-/Planteforsk-ansatte, og det refereres til rett kildehenvisning, f.eks.: "Foto: ... fra Plantevernleksikonet, E. Fløistad, NIBIO".
NIBIO har ikke økonomisk ansvar for tap som måtte oppstå ved bruk av tjenesten.
Plantevernleksikonet © 2026 NIBIO
